Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Ccgsh.tn.edu.tw
Original Paper
Received: November 14, 2001Accepted: December 25, 2001
Natural Plasmid Transformation in Escherichia coli
Suh-Der Tsena Suh-Sen Fanga Mei-Jye Chena Jun-Yi Chiena
aDepartment of Microbiology, Graduate Institute of Microbiology and Immunology, National Yang-Ming Universityand bTaipei American School, Taipei, Taiwan, ROC
Key Words
mids, a most frequent sequence was identified. This
Transformation W Natural transformation, plasmid W
sequence resembled the bacterial interspersed medium
Escherichia coli transformation W Transformation
repetitive sequence of E. coli, suggesting the existence ofa recognition sequence. We conclude that plasmid natu-ral transformation exists in E. coli.Abstract
Copyright 2002 National Science Council, ROC and S. Karger AG, Basel
Although Escherichia coli does not have a natural trans-formation process, strains of E. coli can incorporateextracellular plasmids into cytoplasm ‘naturally’ at low
Among three modes of gene transfer in eubacteria, con-
frequencies. A standard method was developed in which
jugation and transduction can be considered to be the side
stationary phase cells were concentrated, mixed with
effects of plasmid or bacteriophage transmission. In con-
plasmids, and then plated on agar plates with nutrients
trast, transformation might have evolved to incorporate
which allowed cells to grow. Transformed cells could
DNA from the environment [14, 20, 21]. Many genes par-
then be selected by harvesting cells and plating again on
ticipate in this complex process which involves DNA
selective agar plates. Competence developed in the lag
release from donor cells, development of a competent
phase, but disappeared during exponential growth. As
state, recognition of a specific sequence, DNA uptake and
more plasmids were added to the cell suspension, the
DNA integration [20, 21]. The advantage of this process
number of transformants increased, eventually reaching
might be to exchange advantageous properties originating
a plateau. Supercoiled monomeric or linear concatemer-
from separate individuals, to repair damaged DNA, or to
ic DNA could transform cells, while linear monomeric
utilize extracellular DNA as nutrients [13]. These hypoth-
DNA could not. Plasmid transformation was not related
eses are not mutually incompatible, since one process of
to conjugation and was recA-independent. Most of the
DNA uptake might serve several functions. E. coli strains surveyed had this process. All tested plas-
Plasmid is usually transmitted between bacteria by
mids, except pACYC184, could transform E. coli. Inser-
conjugation. Extracellular plasmid is in danger of being
tion of a DNA fragment containing the ampicillin resis-
destroyed. It can get into the cell by exploiting its DNA
tance gene into pACYC184 made the plasmid transform-
uptake process. Since plasmid usually exists in the super-
able. By inserting random 20-base-pair oligonucleotides
coiled form, its entry into the cell and survival inside may
into pACYC184 and selecting for transformable plas-
be difficult if it utilizes the same natural transformation
Graduate Institute of Microbiology and Immunology
National Yang-Ming University, #155, Section 2
Lih-Nong Street, Pei-Tou 11221, Taipei, Taiwan (ROC)
Tel. +886 2 28267108, Fax +886 2 28212880, E-Mail nymutsen@ym.edu.tw
Table 1. Strains and plasmids used
process evolved for linear bacterial DNA. In gram-posi-
tive Bacillus subtilis, plasmid has to be multimeric to be
Initial detection of natural plasmid transformation in starved
cells was done as follows. Stationary phase E. coli K12 cells
transformable [6]. The transformation frequency is much
(CGSC4401) were plated on M9 agar plates. Plasmid p151 (fig. 1b)
lower than that of linear bacterial DNA. These properties
(40 Ìg in 20 Ìl), ampicillin (10 mg/ml, 10 Ìl) and lactose (250 mM,
may reflect both the difficulty of entry and the hostility
20 Ìl) were spotted on this surface in five combinations. After a 12-
that cells have evolved toward an invading foreign genetic
hour incubation at 37 ° C, tiny colonies on plates were replicated on
LB-Ap plates to verify their antibiotic resistance.
Natural transformation has been found in 29 genera of
Transformation Induced by Ampicillin and Cell Lysate
bacteria [14]. In the extensively studied E. coli, natural
Cell lysate was prepared by passing stationary phase CGSC4401
transformation, even though first reported in 1946 by
cells through a French press and centrifuging the resultant lysate at
Boivin [3], has never been confirmed. In this report, we
12,000 rpm for 30 min to sediment intact cells and debris. In the
demonstrate that E. coli can incorporate plasmids without
experiment described in figure 2, stationary E. coli K12 CGSC4401cells were concentrated from 1 ml to 100 Ìl. To this cell suspension
special treatment [12]. Factors influencing plasmid up-
were added glucose (10 Ìl, 250 mM ), plasmid p151 (30 Ìl, 5 Ìg/Ìl)
take and transformability were characterized.
and cell lysate (40 Ìl, equivalent to 108 cells) or ampicillin (20 Ìl, 100 mg/ml). The mixture was then plated on M9 agar and incubated at 37 ° C. After every 2 h, DNase I (2 mg/ml, 40 Ìl) was added to the agar surface and thoroughly mixed with the cells. After another hour Materials and Methods
at 37 ° C, cells were replica-plated on LB-Ap (100 Ìg/ml) plates and incubated at 37 ° C for 16 h to score for transformants. Each point is
Strains and plasmids used are listed in table 1. E. coli strains were
cultured in Luria-Bertani broth or M9 medium supplemented with
Competence Test in Lag Phase, Log Phase, and Stationary Phase
Ca2+ (0.1 mM ), Mg2+ (2 mM ), and glucose (5 mM ) at 37 ° C. When
cells carried antibiotic-resistance markers, the growth medium was
Stationary cells stored at 4 ° C had a lag phase of 9 h when put
supplemented with ampicillin (100 Ìg/ml), kanamycin (50 Ìg/ml) or
back onto fresh M9 glucose plates at 37 ° C. To test lag phase cells for
competence, 2 ! 108 stationary phase cells were mixed with plasmid p151 (2 Ìg), then plated on M9 glucose plates and incubated at 37 ° C Plasmid and Chromosomal DNA Isolation
for 3 h. Plasmids were then digested away with the addition of DNase
Plasmid DNA extraction was carried out using the alkali lysis
I. After 1 h at 37 ° C, cells on M9 glucose plates were collected using
method of Birnboim and Doly [2] as described by Sambrook et al.
M9 medium and washed 3 times in M9 medium. Cells were plated
[16]. Chromosomal DNA was prepared using the method of Silhavy
on LB-Ap plates to score for ampicillin-resistant colonies. To test sta-
et al. [19]. Restriction digestion of purified plasmids and agarose gel
tionary phase cells for competence, cells and plasmids were plated on
(0.8%) electrophoresis was done according to Sambrook et al. [16].
M9 plates without glucose. The rest of the treatment was the same as
Fig. 2. Transformation induced by lysate is 6 h earlier than by ampi- cillin. ) Cells treated with lysate; $ cells treated with ampicillin.
for lag phase cells. To test log phase cells, stationary cells were firstcultured in M9 glucose medium for 9 h, concentrated, mixed withplasmids, and then plated on M9 glucose plates. The rest of the treat-ment was the same as for lag phase cells. Regular Plate AssayAfter knowing that competence was higher in lag phase cells, an
assay procedure was developed. It was the same as the treatment forlag phase cells, except plasmid was not digested using DNase I, butwas washed away using M9 medium. Transformation Using Different E. coli Strains or DifferentPlasmidsWhen testing different strains for transformation by p151, plas-
mid (1.2 mg/ml) was added to 109 3-day-starved cells. Then the regu- lar plate assay was used. When testing pACYC184 plasmid for trans- formability, p151 (1.2 mg/ml), pACYC184 (12 mg/ml) and glucose (25 mM ) were spread on M9 agar with 109 5-day-starved CGSC4401 cells. After 12 h at 37 ° C, cells were replica-plated on either LB-Ap or LB-chloramphenicol plates to score for transformants by either p151 or pACYC184. Fig. 1. Ampicillin and sugar induced starved cells to take up plas-
mid. a Natural transformation on plate. A = Ampicillin; P = plasmid
Restriction endonucleases were purchased from New England
p151; L = lactose. Only cells given lactose, ampicillin and plasmid
Biolabs (Beverly, Mass., USA). Chemicals were purchased from Sig-
DNA gave rise to transformants. This LB-Amp plate was replica-
ma Chemical Co. (St. Louis, Mo., USA). Yeast extract, tryptone and
plated from the original M9 plate which contained starved cells.
agar were purchased from Difco (Detroit, Mich., USA). b Plasmid p151. It contains colE1 replicon, ß-lactamase gene (Apr), ß-galactosidase gene (lacZ) off the Ï PR promoter. The Ï c1857 repressor controlled Ï PR. Plasmids Can ‘Naturally’ Transform E. coli CellsWe explored the possibility of ‘natural’ plasmid trans-
formation using a spot test. Stationary phase E. coli cellswere spread on an M9 minimal salt agar surface without
Fig. 3. Transformants increased as more plasmids were mixed with Fig. 4. Transformation frequency increased as fewer lag phase cells
starved E. coli CGSC4401 cells.
(CGSC4401) were used for transformation.
calcium or magnesium ions. Plasmid, lactose or ampicil-
Although spot tests indicated that cell lysate or ampi-
lin solutions were then spotted on the surface to cover a
cillin was required for plasmid transformation, later de-
small area of cells. After 12 h, cells were inoculated by
tailed experiments revealed that neither was necessary.
replica plating onto LB-Ap agar to select for ampicillin-
Lysate or ampicillin might have augmented the transfor-
resistant cells. Drug-resistant colonies appeared only in
mation frequency so as to allow detection of transfor-
the circle where plasmid, lactose and ampicillin were add-
mants in spot tests. Only glucose or lactose was needed. A
ed. Cells from these colonies were found to contain the
regular plate assay (detailed in Materials and Methods)
plasmid. Thus lactose and ampicillin together seemed to
allow cells to incorporate extracellular plasmid. Plasmidscontained in these colonies were then analyzed using
Competence Higher in Lag Phase Cells
restriction endonucleases. The number and size of restric-
Since starved cells had to be supplied with glucose or
tion fragments of 10 isolates, digested by either EcoRI,
lactose to be transformable, this indicated that starved
BamHI or PstI, were exactly the same as those of the origi-
cells may not be the most receptive ones. To find whether
nal plasmid (data not shown). We then investigated why
there is a ‘competence’ state, plasmids were added to cell
ampicillin was needed to induce transformation in the
cultures in the lag, log or stationary phase for 1 h. Trans-
spot test. Since ampicillin kills most growing cells, we pos-
formants were then scored. Lag phase cells were found to
tulated that lactose allowed some cells to grow and be
be most ‘competent’. The transformation frequency was
killed by ampicillin. The released cell content then stimu-
lower than 10–11/cell for log phase cells, 1.0 ! 10–8/cell
lated surviving cells to incorporate extracellular plasmid
for stationary phase cells, and 1.6 ! 10–5/cell for lag
DNA. If this were true, then cells should be transformable
when supplied with cell lysate in lieu of ampicillin. As
When more plasmids were mixed with cells, more
shown in figure 2, cells given lysate, plasmid and glucose
transformants appeared, but eventually a plateau was
were indeed transformable. The initial appearance of
reached (fig. 3). The linear relationship in the left part of
transformants was earlier in the lysate-treated cells than
figure 3 indicates that one single DNA molecule may
in ampicillin-treated cells (2 vs. 8 h), implying that ampi-
transform one cell. When equal amounts of plasmid DNA
cillin-induced transformants take longer to appear than
were given to increasing numbers of cells, the transforma-
The effect of magnesium and calcium ion on plasmid
Transformation Occurred in Most Tested E. coli
transformation was studied. In standard plate assay, M9
buffer without added Ca2+ or Mg2+ allowed transforma-
Previous experiments employed E. coli strain
tion to happen. When EDTA (2 mM ) was present in the
CGSC4401. Other strains were tested. All strains listed in
M9 (no Ca2+ and Mg2+ added) plate, it totally abolished
table 1 except VGH9761 could be transformed by p151
plasmid transformation. Ca2+ at 0.1 mM allowed transfor-
and pBR322. A comparison of their genotypes indicated
mation, but higher concentrations of Ca2+ did not pro-
that fertility factor F, prophage Ï and the recA gene may
duce higher frequencies of transformation. Increasing
not be critically involved in plasmid transformation.
concentrations of Mg2+ did produce higher frequencies oftransformation, but the highest stimulation at 20 mM of
Untransformable Plasmid Enters Cells when Assisted
Mg2+ was only 15- to 20-fold that of the control experi-
Many kinds of plasmids were tested for transformabili-
ment in which no Mg2+ or Ca2+ was added to M9 buffer.
ty (table 1). Plasmid pBR322 and its derivatives with
We, therefore, supposed that the divalent cation Ca2+ or
inserted E. coli chromosomal DNA could transform E.
Mg2+ played only a permissive function for plasmid natu-
coli. Plasmid pACYC184 did not transform CGSC4401
ral transformation to occur and Mg2+ had a better effect.
cells when given alone or together with p151 in a ratio of
The different effects of Ca2+ and Mg2+ indicated that this
10 (pACYC184) to 1. This amount of p151 generated
plasmid transformation was distinct from the artificial
more than 200 transformant colonies. A mixture of pA-
transformation of plasmid using high concentrations of
CYCl84 with pBR322 did yield natural transformants.
Among the 293 ampicillin-resistant colonies, only 5 colo-
Can transformation occur in liquid suspension? In ten
nies (1.7%) contained both kinds of plasmid. No transfor-
attempts to get transformants by mixing cells and plas-
mant containing pACYC184 alone was found. When
mids in liquid suspension, no transformant formed, indi-
CGSC4401 cells already harboring pBR322 were tested as
cating that E. coli cannot be naturally transformed by
likely recipients, pACYC184 still could not transform
cells. Transformable plasmid seemed to aid untransform-able plasmid to enter cells when both were outside. When
The Effect of Plasmid Structure on Transformation
the former was already inside, it could not help the latter.
In plasmid natural transformation, the structure of the
plasmid affects the transformation activity. We prepared
Some Sequences Helped Plasmid to Transform
three forms of pBR322 plasmid DNA to test their trans-
The inability of pACYC184 to transform cells implies
formability in E. coli. Supercoiled DNA was purified
that E. coli selectively admits plasmids into its cytoplasm.
from agarose gel after electrophoresis. Linear DNA was
Since all plasmids with ampicillin resistance genes could
prepared by digesting supercoiled DNA with restriction
transform, this gene (the 2,208-base pair DNA fragment
endonuclease EcoRI. The concatemeric form was pre-
generated by digesting pACYC177 with restriction en-
pared by ligating linear DNA using T4 DNA ligase. By
zymes BamHI and NheI) was cloned to pACYC184 (a
using the regular plate assay, we found that supercoiled
2,958-base pair DNA fragment generated by digesting
and concatemeric pBR322 could transform E. coli, while
pACYC184 with BamHI and NheI) to test whether any
specific sequence allowed transformation. The resultant
Plasmids linearized separately with either one of two
plasmid pACYC184-177A could naturally transform
restriction endonucleases could still transform E. coli
strains CGSC4401 and CA231. Since this fragment is
when they were mixed together for artificial transforma-
long, a plasmid library was constructed by cloning ran-
tion. In order to know whether similarly treated plasmid
dom 20-base-pair-long oligonucleotides into pACYC184.
could transform naturally, pBR322 was digested with the
This library was used to naturally transform CA231. One
following enzymes separately: AvaI, EcoRI, BamHI, PvuI,
hundred and twenty colonies were picked and the inserted
BsaAI, BsaI, PstI, PvuI. Linearized plasmid was purified
oligonucleotides sequenced. There were 62 kinds of se-
from agarose gel and all eight kinds were mixed together
quences. Those occurring more than once are listed in
in equal amounts. The mixtures were then used to natu-
table 2. One sequence GAAGGAAGATCTTCACCAGT
rally transform E. coli cells. No transformant was ever
occurred 15 times (12.5%) and was called the most fre-
obtained. In control experiments using the artificial trans-
quent sequence (MFS). Although the MFS was not
formation method (calcium chloride treatment and heat
present in the E. coli genome, this MFS shares partial
shock), transformants were obtained.
homology with the palindromic unit (PU) of the bacterial
Fig. 5. Partial homology between transform- able sequences and motifs of BIME. IR = Inverted repeat. Table. 2. Oligonucleotide sequences which made pACYC184 trans-
depends on the presence of a specific DNA sequence on
Can plasmid exploit chromosomal DNA transforma-
tion in order to infect a new host? Plasmid DNA may
have to pass through the machinery evolved for taking up
linear DNA. This poses difficulties for circular or super-
coiled DNA since the uptake process involves binding,
double-stranded DNA cutting, engulfing into transforma-
some, or single-stranded DNA digesting. Plasmid DNA
may be fragmented during this process. This hypothesis
explains why the frequency of plasmid transformation is
much lower than that of chromosomal markers [5, 6, 9,
10, 15, 17, 18, 22]. The inability of monomeric plasmid to
transform and the requirement for a homology region on
the chromosome could also be explained. E. coli does not have a natural transformation process.
The existence of plasmid natural transformation raisesnew questions. Can we hypothesize that DNA transfor-
interspersed mosaic elements (BIME). Another sequence,
mation originally existed but was replaced by conjugation
FS 1, shared partial homology with the inverted-repeat
and transduction and became vestigial? This might ex-
region of the L1 and L2 motifs of the BIME (fig. 5).
plain both the low frequency of transformation and thespecific uptake of certain sequences.
Some E. coli intergenic DNA sequences are composed
Discussion
of repeated BIME. About 500 BIMEs are scattered overthe genome [1, 11]. These BIMEs are composed of a com-
The salient features of this process can be summarized
bination of different conserved motifs. One of these, the
here. Lag phase cells are more receptive to extracellular
PU, was further subdivided into three variants, Y, Z1 and
plasmid than either stationary phase cells or exponential
Z2, differing slightly in sequence. Other motifs include
phase cells. Concatemeric linear, monomeric circular or
seven PU-flanking sequences, called S, L, s, 1, r, A and B.
supercoiled forms of plasmid can transform, whereas
One feature of the PU is the inverted repeats near both
linear monomer cannot. The uptake of plasmid by the cell
ends of the motif. Presumably it can form a stem and loop
structure when the double strands of the PU are opened.
Although BIMEs are found in several enterobacteria,
The MFS which we identified as necessary for plasmid
their functions are unknown. mRNA stabilization, tran-
pACYC184 to transform is partly homologous with the
scription termination, translational control, and genomic
inverted-repeat sequence of PU at 12 nucleotides. FS 1 is
rearrangement were proposed as possible functions. Since
also partly homologous with motifs L1 and L2 (two vari-
each BIME is composed of 2 or more motifs, it may have
eties of L) at 8 nucleotides. These homologies suggest two
several functions simultaneously. The hypothesis that PU
possibilities. First, motifs PU and L in BIME may be the
and L motifs are vestigial recognition sequences for natu-
recognition sequence for E. coli DNA transformation just
ral transformation deserves attention.
like the 11-nucleotide sequence AAGTGCGGTCA forHaemophilus influenzae. The MFS may mimic the recog-nition sequence so as to help untransformable plasmid to
Acknowledgments
transform. Second, the MFS may mimic PU and L motifs
This research was supported by National Science Foundation
in binding to the integration host factor, DNA gyrase or
grant No. NSC892320-B-010-107. We thank Dr. Henry Paulus for
DNA polymerase 1. The binding might protect plasmid
his insightful discussion, Dr. Jon Beckwith, Dr. Mark Krevolin and
from digestion and eventually allow the survival of plas-
Dr. Barbara Bachmann for strains and plasmids. References
1 Bachellier S, Saurin W, Perrin D, Hofnung M,
8 Clowes RC, Hayes W. Experiments in Micro-
15 Mottes M, Grandi G, Sgaramella V, Canosi U,
Gilson E. Structural and functional diversity
bial Genetics. New York, Wiley, 1968.
Morelli G, Trautner TA. Different specific ac-
among bacterial interspersed mosaic elements
9 Contente S, Dubnau D. Characterization of
tivities of the monomeric and oligomeric forms
(BIMEs). Mol Microbiol 12:61–70;1994.
plasmid transformation in Bacillus subtilis: Ki-
of plasmid DNA in transformation of B. subtil-
2 Birnboim HD, Doly J. A rapid alkaline extrac-
netic properties and the effect of DNA confor-
is and E. coli. Mol Gen Genet 1979;174:281–
tion procedure for screening recombinant plas-
mation. Mol Gen Genet 167:251–258;1979.
mid DNA. Nucleic Acids Res 7:1513–1523;
10 De Vos WM, Venema G, Canosi U, Trautner
16 Sambrook J, Fritsch EF, Maniatis T. Molecular
TA. Plasmid transformation in Bacillus subtil-
3 Boivin A. Directed mutation in colon bacilli by
is: Fate of plasmid DNA. Mol Gen Genet 181:
Spring Harbor, Cold Spring Harbor Laborato-
an inducing principle of desoxyribonucleic na-
ture: Its meaning for the general biochemistry
11 Gilson E, Saurin W, Perrin D, Bachellier S,
17 Saunders CW, Guild WR. Monomer plasmid
DNA transforms Streptococcus pneumoniae.
highly repetitive sequences. Res Microbiol 142:
4 Bolivar F, Rodriguez RL, Greene PJ, Betlach
18 Saunders CW, Guild WR. Monomer plasmid
12 Hanahan D. Studies on transformation of Es-
DNA transforms Streptococcus pneumoniae.
and characterization of new cloning vehicles. cherichia coli with plasmids. J Mol Biol 166:
19 Silhavy TJ, Berman ML, Enquist LW. Experi-
5 Canosi U, Iglesias A, Trautner TA. Plasmid
13 Hoelzer MA, Michod RE. DNA repair and the
ments with Gene Fusion. Cold Spring Harbor,
transformation in Bacillus subtilis: Effects of
evolution of transformation in Bacillus subtilis.
Cold Spring Harbor Laboratory Press, 1984.
insertion of Bacillus subtilis DNA into plasmid
20 Smith HO, Danner DB, Deich RA. Genetic
pC 194. Mol Gen Genet 181:434–440;1981.
transformation. Annu Rev Biochem 50:41–68;
6 Canosi U, Morelli G, Trautner TA. The rela-
14 Lorenz MG, Wackernagel W. Bacterial gene
transfer by natural genetic transformation in
21 Stewart GJ, Carlson CA. The biology of natural
transformation efficiency of some S. aureus
the environment. Microbiol Rev 58:563–602;
transformation. Annu Rev Microbiol 40:211–
plasmids isolated from B. subtilis. Mol Gen
22 Stuy JH, Walter RB. Homology-facilitated
plasmid transfer in Haemophilus influenzae.
characterization of amplifiable multicopy
DNA cloning vehicles derived from the P15A
23 Vieira L, Messing J. The pUC plasmids, an
cryptic miniplasmid. J Bacteriol 134:1141–
M13mp7-derived system for insertion muta-
genesis and sequencing with synthetic univer-sal primers. Gene 19:259–268;1982.
2014 Custom Rx TBD for Sheet Metal, Air, Rail, and Transportation Workers - LPPO & RPPO Group Plus Formulary Effective Date: 01/01/2014 - 12/31/2014 Subject to CMS Approval 30 Day Supplies 30 Day Retail from $0 to 30 Day Retail Cost Sharing Out of Pocket that Rx Option Plan/Option Catastrophic (1) from Catastrophic to triggers Unlimited Catastro
PROGRAM FOR KONFERENCEN FM I STRATEGISK LEDELSEVelkomst og indledning Birgitte Dyrvig Carlsson, afdelingsleder, Cowi A/S, ordstyrer, bestyrelsesmedlem, DFM-netværk Facilities Management en strategisk ledelsesdisciplinHovedelementerne i Facilities Management og beskrivelse af, hvordan Facilities Management har Ændring fra udbygningsfokuseret FM til FM som integreret del af museets strategiske
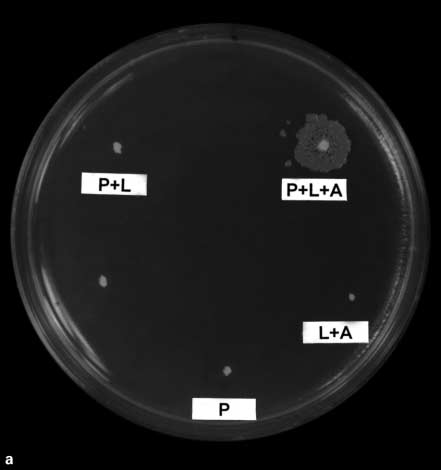 Fig. 2. Transformation induced by lysate is 6 h earlier than by ampi-
Fig. 2. Transformation induced by lysate is 6 h earlier than by ampi-