Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Genetic and expression analyses reveal elevated expression of syntaxin 1a ( stx1a) in high functioning autism
International Journal of Neuropsychopharmacology, Page 1 of 12. Copyright f 2008 CINP
Genetic and expression analyses reveal elevated
expression of syntaxin 1A (STX1A) in highfunctioning autism
Kazuhiko Nakamura1*, Ayyappan Anitha1*, Kazuo Yamada2*, Masatsugu Tsujii3,4,Yoshimi Iwayama2, Eiji Hattori2, Tomoko Toyota2, Shiro Suda1, Noriyoshi Takei1,Yasuhide Iwata1, Katsuaki Suzuki1, Hideo Matsuzaki5, Masayoshi Kawai1,Yoshimoto Sekine1, Kenji J. Tsuchiya1, Gen-ichi Sugihara4, Yasuomi Ouchi4,6,Toshiro Sugiyama7, Takeo Yoshikawa2 and Norio Mori1,4
1 Department of Psychiatry and Neurology, Hamamatsu University School of Medicine,Hamamatsu, Japan2 Laboratory of Molecular Psychiatry, RIKEN Brain Science Institute, Saitama, Japan3 Faculty of Sociology, Chukyo University, Toyota, Aichi, Japan4 The Osaka-Hamamatsu Joint Research, Center for Child Mental Development, Hamamatsu University School of Medicine,Hamamatsu, Japan5 The Osaka-Hamamatsu Joint Research, Center for Child Mental Development, Graduate School of Medicine,Osaka University, Japan6 The Positron Medical Center, Hamamatsu Medical Center, Hamamatsu, Japan7 Aichi Children’s Health and Medical Center, Obu, Aichi, Japan
Autism is a pervasive developmental disorder diagnosed in early childhood. Abnormalities ofserotonergic neurotransmission have been reported in autism. Serotonin transporter (5-HTT), whichmodulates serotonin levels, is a major therapeutic target in autism. Therefore, factors that regulate 5-HTTexpression might be implicated in autism. One candidate 5-HTT-regulatory protein is the presynapticprotein, syntaxin 1A (STX1A). We examined the association of STX1A with autism in a trio associationstudy using DNA samples from 249 AGRE trios with autistic probands. Only male probands were selec-ted, since autism is more prevalent among males. The probands of 102 trios had IQ>70, and were con-sidered as high functioning autism (HFA). In transmission disequilibrium test (TDT) analysis, rs2293485(p=0.034) and rs4717806 (p=0.033) showed nominal associations with HFA ; modest haplotype associ-ation was also observed. The SNPs that showed associations were related to early developmental abnor-malities (ADI-R_D). We further compared STX1A mRNA expression in the lymphocytes of drug-naiveHFA patients (n=12) and age- and sex-matched controls (n=13). STX1A expression in the HFA groupwas significantly higher (p=0.001) than that of controls. Thus, we suggest a possible role of STX1A in thepathogenesis of HFA. During early childhood, there is a period of high brain serotonin synthesis that isdisrupted in autistic children ; STX1A might influence the serotonergic system during this stage of neuro-development, as implied by the association with ADI-R_D.
Received 27 September 2007 ; Reviewed 20 November 2007 ; Revised 9 May 2008 ; Accepted 14 May 2008
Key words : AGRE, high functioning autism, lymphocyte mRNA, serotonin transporter, syntaxin 1A.
communication deficits in combination with restrictedand repetitive behaviours and interests (Kanner, 1943).
Autism is a pervasive developmental disorder diag-
Based on converging evidence from diverse studies,
nosed in early childhood, characterized by social and
the serotonin (5-HT) system can be considered asa logical candidate in autism pathology. The 5-HT
Address for correspondence : K. Nakamura, M.D., Ph.D., Department
system has been found to be developmentally dys-
of Psychiatry and Neurology, Hamamatsu University School of
regulated in autism (Anderson, 1994). 5-HT acts as a
Medicine, Hamamatsu, 431-3192, Japan.
trophic and differentiation factor (Lavdas et al., 1997),
Tel. : +81-53-435-2295 Fax : +81-53-435-3621
in addition to its role as a neurotransmitter. Hyper-
E-mail : nakamura@hama-med.ac.jp* These authors contributed equally to this work.
serotonaemia has been observed in the platelets of
subjects with autism (Ciaranello, 1982 ; Hanley et al.,
lymphocytes (PBL) of drug-naive autistic patients
and control subjects, since lymphocytes are considered
Several studies have reported the association of
as a convenient and accessible alternative to brain
serotonin transporter (5-HTT) polymorphisms with
samples for biochemical and genetic investigations
autism (Betancur et al., 2002 ; Cook et al., 1997 ; Klauck
of the functions of central nervous system (CNS)
et al., 1997 ; Tordjman et al., 2001 ; Yirmiya et al., 2001).
(Gladkevich et al., 2004). We also examined the re-
5-HTT effects the recycling of 5-HT by mediating its
lationship between mRNA expression and genotypes
presynaptic reuptake on termination of serotonergic
neurotransmission ; it is also the major target of selectiveserotonin reuptake inhibitors (SSRIs) (Moore et al.,2004), which have been proven to be efficient in re-
ducing compulsive and stereotyped behaviours and
hyperactivity in autism patients (McDougle et al.,1996). 5-HTT (SLC6A4) is located in 17q11.2, which
has been implicated as an autism linkage region ;
This study was approved by the Ethics Committee of
however, other studies have reported different results
Hamamatsu University School of Medicine. We ob-
with stronger linkage findings (Freitag, 2007). 5-HTT
tained DNA samples from trio families recruited to the
gene promoter variants have been suggested to exert a
Autism Genetic Resource Exchange [AGRE (http://
detectable, albeit small effect on 5-HT blood levels in
www.agre.org) ; Geschwind et al., 2001] ; 249 trio
autistic individuals (Lesch et al., 1996) ; however, as
families with a male proband diagnosed for autism
suggested by Persico et al. (2002) enhanced 5-HT
according to Autism Diagnostic Interview – Revised
blood levels in autism patients, but not in first-degree
(ADI-R ; Lord et al., 1994) and Autism Diagnostic Ob-
relatives with the promoter variant, indicate inter-
servation Schedule (ADOS ; Lord et al., 1989), were
active contributions to hyperserotonaemia both by 5-
selected. Out of the 249 trios, 235 trios were derived
HTT variants and by other autism-related loci. Thus,
from multiplex families ; only male probands [age
factors that regulate the cell surface expression of 5-
9.65¡4.87 yr (mean¡S.D.)] were selected, since autism
HTT might also have a crucial role in the regulation of
has a strong gender bias with a 4 : 1 (male : female)
serotonergic neurotransmission, and therefore, in the
gender ratio. Additional selection criteria required
that (i) there be no possible non-idiopathic autism flag
One candidate 5-HTT-regulatory protein is the pre-
and (ii) all the trios be Caucasian. ADI-R data was
synaptic protein, syntaxin 1A (STX1A) ; this 35-kDa
available for all the 249 affected individuals ; however,
membrane protein belonging to the syntaxin fam-
ADOS testing was available for only 163, and intelli-
ily (Bennett et al., 1992, 1993 ; Inoue et al., 1992) was
gence quotient (IQ) testing for only 112 out of the 249
originally identified as a neuron-specific antigen
autistic individuals. The autistic probands of 102 trios
(Barnstable et al., 1985). Together with the synapto-
had IQ>70, and were considered as the high func-
some-associated protein 25 kDa (SNAP-25) and the
vesicle-associated membrane protein (VAMP), STX1A
We had taken care to exclude the commonly known
forms the soluble N-ethylmaleimide-sensitive factor
genetic causes of autism (e.g. fragile X syndrome), and
attachment protein receptor (SNARE) complex, which
other neurological disorders such as tuberous sclerosis
is vital for chemical neurotransmission (Rothman,
In-vitro studies have shown that STX1A interacts
with 5-HTT, regulating the subcellular localization andexpression of 5-HTT (Haase et al., 2001 ; Quick, 2002).
The genomic structure of STX1A is based on the UCSC
STX1A gene is located at 7q11.23 ; recently, speech de-
May 2004 draft assembly of human genome (http://
lay and autism spectrum behaviours have been found
www.genome.ucsc.edu). STX1A consists of 10 exons
to be associated with duplication of 7q11.23 (Berg et al.,
spanning a genomic stretch of 20.42 kb (mRNA 2064
bases) (Figure 1). In 20 unrelated autism samples selec-
In this trio association study, we examined the SNP
ted randomly from among the AGRE probands, all the
and haplotype associations of STX1A with autism ; we
exons, splice boundaries, and 5k- and 3k-flanking re-
also examined the association of STX1A SNPs with
gions of the gene were screened for polymorphisms by
ADI-R phenotypic data. Further, we compared the
direct sequencing of the respective PCR products.
expression of STX1A mRNA in the peripheral blood
Primer sets used for PCR amplification are listed in
Figure 1. Genomic structure of STX1A and locations of SNPs. Exons are indicated by boxes, with translated regions in closedboxes, and untranslated regions in open boxes ; SNP positions are denoted by arrows.
Table 1. PCR was performed with an initial denatura-
Hardy–Weinberg Equilibrium (HWE) using the
tion at 95 xC for 12 min, followed by 35 cycles at 94 xC
Haploview software version 3.2 (http://www.broad.
for 30 s, 55–60 xC (optimized for each primer pair)
mit.edu/mpg/haploview). Markers were tested for
for 30 s, 72 xC for 1 min, and a final extension at 72 xC
association by transmission disequilibrium test (TDT),
for 7 min, using AmpliTaq Gold DNA polymerase (Ap-
using the TDTPHASE program of the UNPHASED
plied Biosystems, Foster City, CA, USA). Direct se-
software package version 2.403 (http://portal.litbio.
quencing of PCR products was done using the BigDye
org) ; expectation maximization (EM) algorithm was
Terminator Cycle Sequencing kit v3.1 (Applied
used to resolve uncertain haplotypes, to infer missing
Biosystems) in ABI PRISM 3100 Genetic Analyzer
genotypes and to provide maximum-likelihood esti-
Additional SNPs were selected using the infor-
One-way analysis of variance [ANOVA ; GraphPad
mation from International HapMap Project (http://
Prism, version 4.00 (GraphPad Software, San Diego,
www.hapmap.org) and National Centre for Biotech-
CA, USA)] was used to examine the variability in the
nology Information (NCBI dbSNP : http://www.ncbi.
distribution of ADI-R phenotypic data [ADI-R_A (so-
nlm.nih.gov/SNP). On the basis of their genomic
cial interaction), ADI-R_BV (verbal communication),
locations and minor allele frequencies (MAF >0.1)
ADI-R_C (restricted, repetitive and stereotyped pat-
in the Caucasian population, 10 SNPs were chosen
terns of behaviour), ADI-R_D (developmental abnor-
for our analysis, to span the STX1A gene as evenly as
mality prior to 36 months)] across the homozygous
possible. The SNPs used in the study, and their loca-
and heterozygous genotypes of SNPs that showed as-
Linkage disequilibrium (LD) plot was constructed
using the r2 correlation coefficient (Devlin and Risch,
1995) pair-wise LD values between markers were es-
Assay-on-demand/Assay-by-design SNP genotyping
timated using Haploview software. Subsequently,
products (Applied Biosystems) were used to score
associations of haplotypes (frequency >0.01) belong-
SNPs, based on the TaqMan assay method (Ranade
ing to the various haploblocks of STX1A were also
et al., 2001). Genotypes were determined using the
ABI 7900 Sequence Detection System (SDS ; AppliedBiosystems), and analysed using SDS version 2.0 soft-
We obtained blood samples from 12 drug-naive
autistic patients (age 21.4¡2.31 yr) and 13 age-
PedCheck program version 1.1 (http://www.watson.
matched (22.3¡1.93 yr) healthy controls. All the
hgen.pitt.edu) was used to identify and eliminate all
patients and controls were males, and of Japanese
Mendelian inheritance inconsistencies in the trio
origin ; written informed consent was obtained from
genotype data. All the SNPs were tested for
Table 1. PCR primers used for the mutation screening of STX1A gene
The autistic patients were diagnosed according to
sion), respectively. Obsessional/repetitive behaviours
ADI-R (Lord et al., 1994), by trained and certified psy-
were clinically rated using the Yale–Brown Ob-
chiatrists (K.J.T., A.S.). All patients met the autism cri-
sessive–Compulsive Scale (YBOCS ; Goodman et al.,
teria of DSM-IV (APA, 1994) and ICD-10 (WHO, 1992).
1989a, b, Japanese version). Additional aggression
The patients underwent screening, and were excluded
symptoms were also assessed using the aggression
if they had any major medical or psychiatric conditions.
questionnaire (AQ ; Buss and Perry, 1992, translation
We had taken care to exclude the commonly known
in Japanese). A faux pas detection task was used to
genetic causes of autism (e.g. fragile X syndrome), and
measure theory of mind (Baron-Cohen et al., 1999,
other neurological disorders such as tuberous sclerosis
Stone et al., 2003, translation in Japanese). All the
and neurofibromatosis. All the patients were of the
evaluations were conducted by a trained research
HFA type ; the average IQ scores were measured by the
Wechsler Adult Intelligence Scale – Revised (WAIS-R ;
All the controls were free of medications, and under-
went screening to exclude neurological, developmen-
We used the following instruments for the assess-
tal, or psychiatric disorders and mental retardation ;
ment of autistic symptoms. Comorbid anxiety and
none of them met any of the relevant criteria of
depressive symptoms were assessed using the
DSM-IV. The average IQ of the control subjects (meas-
Hamilton Anxiety Rating Scale (HAMA ; Hamilton,
ured by WAIS-R, Wechsler, 1981, Japanese version)
1959, Japanese version) and the Hamilton Depression
did not differ significantly from that of the HFA pa-
Rating Scale (HAMD ; Hamilton, 1960, Japanese ver-
Table 2. Single SNP transmission disequilibrium test (TDT) results of STX1A SNPs
HFA, High functioning autism ; T, transmitted ; UTR, untranslated region. a Common allele is listed first. b Based on the parental genotypes of 249 trios. c T % of common allele is listed. d Computed on the basis of likelihood ratio test ; significant p values (<0.05) are indicated in bold italics. e 10 000 permutations.
(GAPDH) that served as the endogenous reference,were purchased from Applied Biosystems (Assay-on-
Peripheral blood (20 ml) was drawn from the cubital
DemandTM gene expression products Hs00195648 and
vein into EDTA-containing plastic syringes. Lym-
Hs99999905, respectively). All reactions were per-
phocytes were isolated from blood samples by the
formed in duplicate, according to the manufacturer’s
Ficoll-Paque gradient method, and total RNA was
protocol. A comparative threshold cycle (C
extracted using RNAzolB reagent (Sawady, Tokyo,
validation experiment was performed to check if the
Japan) according to the manufacturer’s instructions.
efficiencies of target and reference amplifications were
RNA samples were quantified by analysing the
approximately equal (the slope of the log input amount
absorbance at 260 nm in a UV spectrophotometer.
Complementary DNA (cDNA) was synthesized by
T<0.1). One sample was randomly chosen as the
calibrator and was amplified in each plate to correct for
first-strand reverse transcriptase (RT) reaction using
the experimental differences among consecutive PCR
Random Primer and M-MLV reverse transcriptase
runs. The amounts of STX1A mRNA were normalized
to the endogenous reference and expressed relative
We had maintained similar conditions during the
to the calibrator as 2xDDCT (comparative C
collection and processing of all the samples since cir-
cadian regulation of STX1A has been reported(Allaman-Pillet et al., 2004). The blood samples were
collected at around 10:00 hours on the days assignedfor sample collection. Lymphocyte isolation and RNA
Statistical calculations were performed using SPSS
extraction were done immediately thereafter, under
statistical package, version 11.0.1 (SPSS Co. Ltd,
Tokyo, Japan) and GraphPad Prism. The differencein STX1A expression between groups was analysedusing t test. Correlation between the various clinical
Quantitative reverse transcriptase polymerase chain
features of HFA group and STX1A expression was
examined by Spearman’s rank correlation test. Fur-
Real-time qRT–PCR analysis was performed using
ther, regression analysis was carried out to check
ABI PRISM 7900 Sequence Detection System (Ap-
the effect of comorbidities, including obsession-
plied Biosystems). TaqMan primer/probes for STX1A
compulsion (YBOCS scores) and aggression, on
and for glyceraldehyde-3-phosphate dehydrogenase
Figure 2. Comparison of the distribution of ADI-R_D scores across the homozygous and heterozygous genotypes of(a) SNP08 (p=0.036), (b) SNP03 (p=0.04), (c) SNP09 (p=0.036), (d) SNP10 (p=0.029), in the probands of high functioningautism (HFA) trios.
Relationship between mRNA expression and genotypes
results are shown in Table 2. In the TDT of HFA trios,SNP04 (p=0.034, OR 1.54, 95 % CI 1.03–2.29) and
All the samples from the mRNA expression study
SNP08 (p=0.033, OR 0.63, 95 % CI 0.41–0.96) showed
were genotyped for the SNPs that showed associations
nominal associations. Overtransmission was observed
in the trio association study. Using the Kruskal–Wallis
for the minor allele T (56.07 %) of SNP04 (rs941298 ;
test, we examined the relationship between genotypes
exon 3, D68D) and for the minor allele A (57.69 %) of
SNP08 (rs4717806). SNP03 (p=0.054), SNP05 (p=0.053), SNP09 (p=0.050) and SNP10 (p=0.050)
showed tendencies for association with HFA. The
global p value, however, was not significant (p=0.124).
None of the SNPs showed significant association in
No new mutations were observed in the coding orregulatory regions of the gene, other than the SNPs
We examined the associations of ADI-R phenotypicdata (ADI-R_A, ADI-R_BV, ADI-R_C, and ADI-R_D)
with the SNPs that showed nominal associations in
Mendelian inheritance inconsistencies were not ob-
single SNP TDT. In the HFA trios, significant associ-
served for any of the SNPs. More than 98 % of the
ation with ADI-R_D scores was observed for SNP08
genotypes were scored for each SNP ; none of the SNPs
(p=0.036) ; ADI-R_D was lower in the A/A group of
this SNP (Figure 2a). We also examined the associ-
TDT was performed separately for the whole set of
ations of SNPs that showed tendency for association in
249 trios and for the HFA subgroup of 102 trios ; the
single SNP TDT. SNP03 (p=0.040), SNP09 (p=0.036)
correlation was observed between any of the clinical
features of HFA group and STX1A expression
(Table 4). Further, regression analysis showed that
there is no effect of YBOCS/aggression on STX1A
By Kruskal–Wallis test, the relationship between geno-
types and STX1A mRNA expression was examined
for SNP04 and SNP08, which showed nominal asso-
Figure 3. Haplotype block structure of STX1A based on r2
ciations with HFA in TDT analysis. SNP08 showed a
values calculated from 102 high functioning autism (HFA)
tendency for gene dose effect, with the following
trend : expression of A/A genotype (mean¡S.D.)(0.243¡0.162)<expression of A/T genotype (0.293¡0.143)<expression of T/T genotype (0.462¡0.023) ;
and SNP10 (p=0.029) showed significant associations
however, the difference was not statistically signifi-
with ADI-R_D ; the scores were lower in the A/A
group of SNP03, in the C/C group of SNP09 and in theC/C group of SNP10 (Figure 2b–d, respectively).
After adjusting for IQ effects with univariate ana-
lysis of variance, there were only tendencies for
In this study, STX1A mRNA expression was signifi-
association with ADI-R_D (SNP08, p=0.070 ; SNP03,
cantly higher in the lymphocytes of drug-naive HFA
p=0.070 ; SNP09, p=0.070 ; SNP10, p=0.055).
patients compared with controls. Lymphocytes are
No associations were observed with other pheno-
considered as excellent neural probes for studying
typic data. In the whole group of 249 trios, there were
neuropsychiatric disorders due to (i) altered lympho-
no significant associations with any of the ADI-R
cyte functions in neuropsychiatric conditions, (ii) ex-
pression of neuroactive proteins and processes inlymphocytes, and (iii) similarities of hormonal effects
on the nervous system processes and lymphocyte
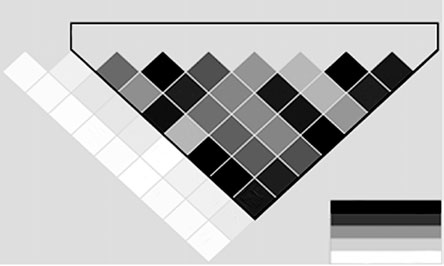
LD analysis identified a single haploblock across the
physiology (reviewed by Gladkevich et al., 2004). In
STX1A gene, comprising SNP03 to SNP10 (Figure 3).
the present study, there was no significant correlation
The LD pattern observed in our study is similar to that
between STX1A expression and any of the clinical
reported for the 30 CEU (CEPH) trios in the HapMap
features of the HFA group ; therefore, altered STX1A
expression may be implicated in the general patho-physiology of HFA, rather than in the symptoms of the
disease. The possibilities of using the enhanced ex-pression of STX1A as a peripheral marker in the di-
The results of haplotype association analysis are
agnosis of HFA may be explored. However, our study
shown in Table 3. The haplotype ATACCTCC showed
is limited by small sample size ; therefore, replication
a nominal association in the HFA subgroup of 102
in a larger sample size is warranted.
trios (p=0.044) ; however, this was not significant after
Lymphoblastoid cells lines have been reported as
the permutation test (10 000 permutations). None of
viable tools for identifying the genes associated with
the haplotypes showed any significant association in
autism (Baron et al., 2006 ; Hu et al., 2006 ; Nishimura
et al., 2007). There is a close association between thestate of the immune system, particularly lymphocytes,
and major psychiatric disorders including autism
Descriptive clinical data of the autism samples is pro-
(Ashwood et al., 2006 ; Krause et al., 2002). STX1A is a
vided in Table 4. STX1A expression in the drug-naive
neuron-specific antigen (Barnstable et al., 1985),
HFA group (0.38¡0.39) was found to be significantly
which, in association with SNAP-25 and VAMP, forms
higher (t=x4.37, d.f.=14, p=0.001) than that of the
the SNARE complex that is vital for chemical neuro-
control group (0.17¡0.06) (Figure 4). No significant
transmission (Rothman, 1994 ; Sollner et al., 1993). In a
Table 3. Haplotype associations of SNPs belonging to the single LD block of STX1A in102 HFA trios
LD, Linkage disequilibrium ; HFA, high functioning autism. a All possible combinations of haplotypes with frequency >0.01. b Significant p values (<0.05) are indicated in bold italics. c 10 000 permutations.
Table 4. Descriptive data on autism samples and their correlation with STX1A mRNAexpression
HAMD, Hamilton Depression Rating Scale ; HAMA, Hamilton Anxiety Rating Scale ;YBOCS, Yale–Brown Obsessive–Compulsive Scale.
process similar to the exocytosis of neurotransmitters,
comparable to the STX1A expression during their first
STX1A and other SNARE proteins are involved in a
years of life, which is the most critical period for the
process of exocytosis of lytic granules, mediating tar-
formation of neural wiring patterns.
get cell death in the PBLs that play a pivotal role in the
In the trio association study, nominal SNP and
body’s defence against infection (Hong, 2005). The
haplotype associations were observed with HFA.
presence of sympathetic fibres in lymphoid tissues
None of the SNP and haplotype associations will re-
suggests a direct contact area for neural signalling
main significant after multiple testing correction ;
cascade with the immune cells (Felten et al., 1987).
however, it is difficult to apply a Bonferroni correction
Thus, lymphocyte STX1A expression might be indi-
in this study, since strong LD was observed between
cative of CNS expression. In animal models, STX1A
SNP03 and SNP10. Further, single genes are likely to
expression has been shown to be more or less constant
have only small individual effects in complex dis-
after the embryonic and immediate postnatal stages
orders like autism. Among the two SNPs that showed
(Biranowska et al., 2002 ; Shimohama et al., 1998).
nominal associations with HFA, rs2293485 is a syn-
Therefore, the expression pattern of STX1A in our
onymous exonic SNP, while rs4717806 is an intronic
study subjects with an average age of 22 yr might be
SNP. The other SNPs that showed tendencies for
should precede the appearance of clinical symptoms.
The importance of 5-HT in several aspects of prenataland postnatal brain development that are related to
autism is well documented. Humans undergo a periodof high brain 5-HT synthesis capacity during early
childhood, which is disrupted in autistic children
(Chugani et al., 1999). Thus, STX1A might be involvedin the pathogenesis of autism by influencing the sero-
tonergic system, during the early stages of brain de-
velopment. However, after adjustment for IQ, there
were only tendencies for association with ADI-R_D.
There was no relation with other ADI-R subscores
(ADI-R_A, ADI-R_BV, ADI-R_C), which reflect the
During the postnatal stages of neurodevelopment,
the expression of 5-HTT might be a prerequisite for the
neurodevelopmental functions exerted by 5-HT. In-
Figure 4. t test comparison of STX1A mRNA levels in the
vitro studies have shown that 5-HTT expression is
lymphocytes from control subjects and drug-naive high
regulated by STX1A (Haase et al., 2001 ; Quick, 2002).
functioning autism (HFA) patients. Expression level of
In cultured cells, STX1A has been found to interact
STX1A is normalized against that of glyceraldehyde-3-
with 5-HTT, modulating the cell-surface expression of
phosphate dehydrogenase (GAPDH). Horizontal bars
5-HTT (Haase et al., 2001). Cells transfected with
indicate means. A significant difference in STX1A expression
STX1A cDNA showed a decrease in the number of 5-
was observed between the two groups (p=0.001).
HTT molecules expressed on the cell surface ; the de-crease in 5-HTT expression was suggested to be
association are also intronic SNPs. Therefore, the as-
caused either by an inhibition of the recycling of pre-
sociated SNPs may not have a direct influence on
viously internalized 5-HTT or by a blocking of the
STX1A expression. It is possible that the expression is
trafficking of newly synthesized 5-HTT to the plasma
regulated by additional trans-acting factors in a net-
membrane. Studies using thalamocortical neuronal
cultures also have shown that STX1A affects the loca-
Considering sample homogeneity, we had selected
lization and cell-surface expression of 5-HTT (Quick,
predominantly high-functioning trio samples, where
2002). Thus, STX1A might be suggested to influence
all the subjects were strictly diagnosed with autistic
the serotonergic system by modulating the expression
disorder. Since mental retardation consists of various
aetiologies, HFA samples can be considered to be
An alternate mode of action of STX1A may be
relatively free from non-specific environmental or
through its interaction with the glutamatergic system
genetic risks for their impairments. We had used all
(Fan et al., 2006 ; Yu et al., 2006). Glutamate, the major
available AGRE cases that satisfied our strict selection
excitatory neurotransmitter in the brain, plays a vital
criteria at the time of subject selection.
role in brain development, affecting neuronal mi-
We observed an association of ADI-R_D scores with
gration, neuronal differentiation, axon genesis, and
the SNPs that showed nominal associations or ten-
neuronal survival (Coyle et al., 2002). STX1A promotes
dencies for association with HFA. This observation in
the endocytic sorting of the glutamate transporter
HFA patients who had been diagnosed for autism is
EAAC1, leading to inhibition of glutamate transport
interesting, since ADI-R_D reflects developmental ab-
(Yu et al., 2006). Several studies have implicated ab-
normalities observed in autism prior to 36 months ;
normalities of glutamatergic neurotransmission in the
delayed speech, unusual socio-emotional reactions
pathophysiology of autism (reviewed by McDougle et
and poor attentions to and exploration of the en-
al., 2005). Glutamate-related genes have also been
vironment, are among the first clinically noticeable
shown to be important in the pathology of autism
behavioural symptoms of autism during the second
and third years of life (Dahlgren and Gillberg, 1989 ;
In conclusion, we suggest a possible role of STX1A
De Giacomo and Fombonne, 1998). Since autism is
in the pathogenesis of HFA ; this is the first report of an
a pervasive developmental disorder, neuroanatomi-
association between STX1A and HFA. In autism,
cal, neurochemical and neurobiological abnormalities
mental retardation implies the involvement of more
risk factors and genes ; this might be the reason that
Bennett MK, Calakos N, Scheller RH (1992). Syntaxin : a
the autism group, as a whole, was not associated with
synaptic protein implicated in docking of synaptic vesicles
STX1A. The association of STX1A with the HFA group
at presynaptic active zones. Science 257, 255–259.
may thus, indicate a protective role against mental re-
Bennett MK, Garcia-Arraras JE, Elferink LA, Peterson K,
Fleming AM, Hazuka CD, Scheller RH (1993). The
tardation. However, considering our sample size, it
syntaxin family of vesicular transport receptors. Cell 74,
may be too premature to draw such a conclusion ; the
elevated expression of STX1A might also be secondary
Berg JS, Brunetti-Pierri N, Peters SU, Kang SH, Fong CT,
to other metabolic changes in HFA. Therefore, further
Salamone J, Freedenberg D, Hannig VL, Prock LA, Miller
studies into the functional impact of STX1A in HFA
DT, et al. (2007). Speech delay and autism spectrum
behaviors are frequently associated with duplication of the7q11.23 Williams-Beuren syndrome region. Genetics inMedicine 9, 427–441.
Betancur C, Corbex M, Spielewoy C, Philippe A, Laplanche
We gratefully acknowledge the support of the Autism
JL, Launay JM, Gillberg C, Mouren-Simeoni MC, Hamon
Genetics Resource Exchange (AGRE, www.agre.org).
M, Giros B, Nosten-Bertrand M, Leboyer M (2002). Serotonin transporter gene polymorphisms and
This work was supported by Research on Brain
hyperserotonemia in autistic disorder. Molecular Psychiatry
Science Funds from the Ministry of Health Labor and
Welfare, Japan and a Grant-in-Aid for Scientific Re-
Biranowska J, Dziewiatkowski J, Ludkiewicz B, Morys J
search (C) from the Ministry of Education, Culture,
(2002). Developmental changes of synaptic proteins
Sports, Science and Technology of Japan. We thank
expression within the hippocampal formation of the rat.
Hosoya Teruyo for technical assistance.
Anatomy and Embryology 206, 85–96.
Buss AH, Perry M (1992). The aggression questionnaire.
Journal of Personality & Social Psychology 63, 452–459.
Chugani DC, Muzik O, Behen M, Rothermel R, Janisse JJ,
Lee J, Chugani HT (1999). Developmental changes in brain
serotonin synthesis capacity in autistic and nonautisticchildren. Annals of Neurology 45, 287–295.
Ciaranello RD (1982). Hyperserotonemia and early
infantile autism. New England Journal of Medicine 307,
Allaman-Pillet N, Roduit R, Oberson A, Abdelli S, Ruiz J,
Beckmann JS, Schorderet DF, Bonny C (2004). Circadian
Cook Jr. EH, Courchesne R, Lord C, Cox NJ, Yan S, Lincoln
regulation of islet genes involved in insulin production
A, Haas R, Courchesne E, Leventhal BL (1997). Evidence
and secretion. Molecular and Cellular Endocrinology 226,
of linkage between the serotonin transporter and autistic
disorder. Molecular Psychiatry 2, 247–250.
Anderson GM (1994). Studies on the neurochemistry of
Coyle JT, Leski ML, Morrison JH (2002). The diverse roles
autism. In : Bauman ML, Kemper TL (Eds), The
of l-glutamic acid in brain signal transduction. In :
Neurobiology of Autism (pp. 227–242) Baltimore : Johns
Davis KL, Charney D, Coyle JT, Nemeroff C (Eds),
Neuropsychopharmacology : The Fifth Generation of Progress
APA (1994). Diagnostic and Statistical Manual of Mental
(pp. 71–90). Philadelphia : Lippincott Williams & Wilkins.
Disorders (4th edn). Washington DC : American Psychiatric
Dahlgren SO, Gillberg C (1989). Symptoms in the first two
years of life. A preliminary population study of infantile
Ashwood P, Wills S, Van de Water J (2006). The immune
autism. European Archives of Psychiatry and Neurological
response in autism : a new frontier for autism research.
Journal of Leukocyte Biology 80, 1–15.
De Giacomo A, Fombonne E (1998). Parental recognition of
Barnstable CJ, Hofstein R, Akagawa K (1985). A marker of
developmental abnormalities in autism. European Child and
early amacrine cell development in rat retina. Brain
Depienne C, Heron D, Betancur C, Benyahia B, Trouillard
Baron CA, Liu SY, Hicks C, Gregg JP (2006). Utilization of
O, Bouteiller D, Verloes A, LeGuern E, Leboyer M, Brice
lymphoblastoid cell lines as a system for the molecular
A (2007). Autism, language delay and mental retardation
modeling of autism. Journal of Autism and Developmental
in a patient with 7q11 duplication. Journal of Medical
Baron-Cohen S, O’Riordan M, Stone V, Jones R, Plaisted K
Devlin B, Risch N (1995). A comparison of linkage
(1999). Recognition of faux pas by normally developing
disequilibrium measures for fine-scale mapping. Genomics
children and children with Asperger syndrome or
high-functioning autism. Journal of Autism and
Fan HP, Fan FJ, Bao L, Pei G (2006). SNAP-25/syntaxin 1A
Developmental Disorders 29, 407–418.
complex functionally modulates neurotransmitter
gamma-aminobutyric acid reuptake. Journal of Biological
Lavdas AA, Blue ME, Lincoln J, Parnavelas JG (1997).
Serotonin promotes the differentiation of glutamate
Felten DL, Felten SY, Bellinger DL, Carlson SL, Ackerman
neurons in organotypic slice cultures of the developing
KD, Madden KS, Olschowki JA, Livnat S (1987).
cerebral cortex. Journal of Neuroscience 17, 7872–7880.
Noradrenergic sympathetic neural interactions with the
Lesch KP, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri
immune system : structure and function. Immunological
S, Benjamin J, Muller CR, Hamer DH, Murphy DL (1996).
Association of anxiety-related traits with a polymorphism
Freitag CM (2007). The genetics of autistic disorders and its
in the serotonin transporter gene regulatory region. Science
clinical relevance : a review of the literature. Molecular
Lord C, Rutter M, Goode S, Heemsbergen J, Jordan H,
Geschwind DH, Sowinski J, Lord C, Iversen P, Shestack J,
Mawhood L, Schopler E (1989). Autism diagnostic
Jones P, Ducat L, Spence SJ (2001). The autism genetic
observation schedule : a standardized observation of
resource exchange : a resource for the study of autism and
communicative and social behavior. Journal of Autism and
related neuropsychiatric conditions. American Journal of
Developmental Disorders 19, 185–212.
Lord C, Rutter M, Le Couteur A (1994). Autism Diagnostic
Gladkevich A, Kauffman HF, Korf J (2004). Lymphocytes
Interview – Revised : a revised version of a diagnostic
as a neural probe : potential for studying psychiatric
interview for caregivers of individuals with possible
disorders. Progress in Neuropsychopharmacology and
pervasive developmental disorders. Journal of Autism and
Biological Psychiatry 28, 559–576.
Developmental Disorders 24, 659–685.
Goodman WK, Price LH, Rasmussen SA, Mazure C,
McDougle CJ, Erickson CA, Stigler KA, Posey DJ (2005).
Delgado P, Heninger GR, Charney DS (1989a). The
Neurochemistry in the pathophysiology of autism.
Yale-Brown Obsessive Compulsive Scale. II. Validity.
Journal of Clinical Psychiatry 66 (Suppl. 10), 9–18.
Archives of General Psychiatry 46, 1012–1016.
McDougle CJ, Naylor ST, Cohen DJ, Volkmar FR,
Goodman WK, Price LH, Rasmussen SA, Mazure C,
Heninger GR, Price LH (1996). A double-blind,
Fleischmann RL, Hill CL, Heninger GR, Charney DS
placebo-controlled study of fluvoxamine in adults with
(1989b). The Yale-Brown Obsessive Compulsive Scale.
autistic disorder. Archives of General Psychiatry 53,
I. Development, use, and reliability. Archives of General
Moore ML, Eichner SF, Jones JR (2004). Treating functional
Haase J, Killian AM, Magnani F, Williams C (2001).
impairment of autism with selective serotonin-reuptake
Regulation of the serotonin transporter by interacting
inhibitors. Annals of Pharmacotherapy 38, 1515–1519.
proteins. Biochemical Society Transactions 29, 722–728.
Nishimura Y, Martin CL, Vazquez-Lopez A, Spence SJ,
Hamilton M (1959). The assessment of anxiety states by
Alvarez-Retuerto AI, Sigman M, Steindler C, Pellegrini S,
rating. British Journal of Medical Psychology 32, 50–55.
Schanen NC, Warren ST, Geschwind DH (2007).
Hamilton M (1960). A rating scale for depression. Journal of
Genome-wide expression profiling of lymphoblastoid
Neurology, Neurosurgery and Psychiatry 23, 56–62.
cell lines distinguishes different forms of autism and
Hanley HG, Stahl SM, Freedman DX (1977).
reveals shared pathways. Human Molecular Genetics 16,
Hyperserotonemia and amine metabolites in autistic and
retarded children. Archives of General Psychiatry 34,
Persico AM, Pascucci T, Puglisi-Allegra S, Militerni R,
Bravaccio C, Schneider C, Melmed R, Trillo S,
Hong W (2005). Cytotoxic T lymphocyte exocytosis : bring on
Montecchi F, Palermo M, et al. (2002). Serotonin
the SNAREs! Trends in Cell Biology 15, 644–650.
transporter gene promoter variants do not explain the
Hu VW, Frank BC, Heine S, Lee NH, Quackenbush J (2006).
hyperserotoninemia in autistic children. Molecular
Gene expression profiling of lymphoblastoid cell lines
from monozygotic twins discordant in severity of autism
Quick MW (2002). Role of syntaxin 1A on serotonin
reveals differential regulation of neurologically relevant
transporter expression in developing thalamocortical
neurons. International Journal of Developmental Neuroscience
Inoue A, Obata K, Akagawa K (1992). Cloning and sequence
analysis of cDNA for a neuronal cell membrane antigen,
Ranade K, Chang MS, Ting CT, Pei D, Hsiao CF, Olivier M,
HPC-1. Journal of Biological Chemistry 267, 10613–10619.
Pesich R, Hebert J, Chen YD, Dzau VJ, et al. (2001).
Kanner L (1943). Autistic disturbances of affective contact.
High-throughput genotyping with single nucleotide
polymorphisms. Genome Research 11, 1262–1268.
Klauck SM, Poustka F, Benner A, Lesch KP, Poustka A
Rothman JE (1994). Mechanisms of intracellular protein
(1997). Serotonin transporter (5-HTT) gene variants
associated with autism? Human Molecular Genetics 6,
Shimohama S, Fujimoto S, Sumida Y, Akagawa K, Shirao T,
Matsuoka Y, Taniguchi T (1998). Differential expression of
Krause I, He XS, Gershwin ME, Shoenfeld Y (2002). Brief
rat brain synaptic proteins in development and aging.
report : immune factors in autism : a critical review. Journal
Biochemical and Biophysical Research Communications 251,
of Autism and Developmental Disorders 32, 337–345.
Sollner T, Bennett MK, Whiteheart SW, Scheller RH,
gene in the behavioral expression of autism. Molecular
Rothman JE (1993). A protein assembly-disassembly
pathway in vitro that may correspond to sequential steps
Wechsler D (1981). Wechsler Adult Intelligence Scale –
of synaptic vesicle docking, activation, and fusion.
Revised (WAIS-R). New York : The Psychiatric
Stone VE, Baron-Cohen S, Calder A, Keane J, Young A
WHO (1992). The ICD-10 Classification of Mental and Behavioral
(2003). Acquired theory of mind impairments in
Disorders : Diagnostic criteria for research. Geneva : World
individuals with bilateral amygdala lesions.
Yirmiya N, Pilowsky T, Nemanov L, Arbelle S, Feinsilver T,
Szatmari P, Paterson AD, Zwaigenbaum L, Roberts W,
Fried I, Ebstein RP (2001). Evidence for an association
Brian J, Liu XQ, Vincent JB, Skaug JL, Thompson AP,
with the serotonin transporter promoter region
Senman L, et al. (2007). Mapping autism risk loci using
polymorphism and autism. American Journal of Medical
genetic linkage and chromosomal rearrangements. Nature
Yu YX, Shen L, Xia P, Tang YW, Bao L, Pei G (2006).
Tordjman S, Gutknecht L, Carlier M, Spitz E, Antoine C,
Syntaxin 1A promotes the endocytic sorting of EAAC1
Slama F, Carsalade V, Cohen DJ, Ferrari P, Roubertoux
leading to inhibition of glutamate transport. Journal of Cell
PL, Anderson GM (2001). Role of the serotonin transporter
Tamoxifen therapy in breast cancer control worldwide. Richard R. Love1 and Valentin Koroltchouk21Professor, Departments of Human Oncology, Medicine, and Family Medicine and Practice,University of Wisconsin - Madison. Mailing address: 7C, 1300 University Avenue, Madison,WI 53706, USA2Scientist, Cancer and Palliative Care Unit, World Health Organization, Geneva, Switzerland. Mailing address: Wo
Nº 105, quinta-feira, 31 de maio de 2012Metronidazol, 100MG/G creme vaginal Prati DonaduzziEnvelope Sais para reidratação oral, envelope con- Prati DonaduzziBisnaga Sulfadiazina de Prata 1%, Creme c/ Prati DonaduzziOBSERVAÇÃO: A descrição completa do produto encontra-se disponível na Secretaria Municipal de DESPACHOS DO PREFEITO AVISO DE LICITAÇÃO(*) EXTRATO DO CONTRATO No-
 correlation was observed between any of the clinical
features of HFA group and STX1A expression
(Table 4). Further, regression analysis showed that
there is no effect of YBOCS/aggression on STX1A
By Kruskal–Wallis test, the relationship between geno-
types and STX1A mRNA expression was examined
for SNP04 and SNP08, which showed nominal asso-
Figure 3. Haplotype block structure of STX1A based on r2
ciations with HFA in TDT analysis. SNP08 showed a
values calculated from 102 high functioning autism (HFA)
tendency for gene dose effect, with the following
trend : expression of A/A genotype (mean¡S.D.)(0.243¡0.162)<expression of A/T genotype (0.293¡0.143)<expression of T/T genotype (0.462¡0.023) ;
and SNP10 (p=0.029) showed significant associations
however, the difference was not statistically signifi-
with ADI-R_D ; the scores were lower in the A/A
group of SNP03, in the C/C group of SNP09 and in theC/C group of SNP10 (Figure 2b–d, respectively).
correlation was observed between any of the clinical
features of HFA group and STX1A expression
(Table 4). Further, regression analysis showed that
there is no effect of YBOCS/aggression on STX1A
By Kruskal–Wallis test, the relationship between geno-
types and STX1A mRNA expression was examined
for SNP04 and SNP08, which showed nominal asso-
Figure 3. Haplotype block structure of STX1A based on r2
ciations with HFA in TDT analysis. SNP08 showed a
values calculated from 102 high functioning autism (HFA)
tendency for gene dose effect, with the following
trend : expression of A/A genotype (mean¡S.D.)(0.243¡0.162)<expression of A/T genotype (0.293¡0.143)<expression of T/T genotype (0.462¡0.023) ;
and SNP10 (p=0.029) showed significant associations
however, the difference was not statistically signifi-
with ADI-R_D ; the scores were lower in the A/A
group of SNP03, in the C/C group of SNP09 and in theC/C group of SNP10 (Figure 2b–d, respectively).