Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Pii: s0304-3940(98)00302-4
Neuroscience Letters 247 (1998) 147–150
Characterization of a plasma membrane zinc transporter in rat brain
Department of Biological Sciences, Program in Neurobiology, Ohio University, Athens OH 45701, USA
Received 19 February 1998; received in revised form 26 March 1998; accepted 31 March 1998
Abstract
Many studies now show that zinc plays a critical and unique role in central nervous system development and function. The
cellular mechanisms of zinc efflux and influx are largely unknown and few models exist that describe cellular zinc transport in thebrain. This report provides convincing evidence of a zinc transporter in plasma membrane vesicles isolated from rat brain. Zincuptake was saturable ( K =
10 nmol/mg per 30 s), was seen in the absence of ATP, and was unaffected by
gradients for other ions such as Na+ or K+. Increasing the ionic strength of the extravesicular media with Na+, K+, or choline+reduced zinc uptake approximately 50%. Whereas, increasing extravesicular H+ concentration (pH = 5) resulted in near com-plete inhibition of zinc uptake. Intravesicular zinc was rapidly released upon lowering extravesicular concentrations of zinc withthe heavy metal chelator O-phenanthroline (1 mM). The results are consistent with a freely-reversible transport of zinc across theplasma membrane of neurons. 1998 Elsevier Science Ireland Ltd.
Keywords: Ion transport; Membrane vesicles; Excitotoxicity; Zinc homeostasis; Transition metals; Alzheimer’s disease
Zinc levels and localization in the brain are tightly regu-
tissues and cell types (for reviews see [4,11]). These studies
lated. As much as 15% of brain zinc is sequestered in synap-
have resulted in the cloning of three mammalian zinc trans-
tic vesicles [3] associated with excitatory amino acid
porters [9,10]. ZnT-1 is associated with the plasma mem-
neurotransmitters. This is particularly true of mossy fiber
brane; ZnT-2 is associated with the endoplasmic reticulum,
terminals in the hippocampus. Stimulation of the mossy
and ZnT-3 is associated with synaptic vesicles in hippocam-
fiber pathway in the hippocampus results in release and
pus and cortex. However, the mechanism of transport for
trans-synaptic movement of zinc [2]. The vast majority of
these three zinc transporters is poorly understood, although
intracellular zinc is tightly bound to zinc binding proteins
they show specificity for zinc. A fourth type of metal ion
such as metallothionien III, which is specifically expressed
transporter has been cloned from rat intestine and is termed
in brain [5]. Understanding how cellular zinc homeostasis is
DCT1 [6]. This transporter is quite different from the ZnT
maintained in the brain is essential, as dysregulation of zinc
family and is highly expressed in the proximal duodenum,
levels is potentially very neurotoxic [2]. Neuronal zinc
kidney, thymus, and brain. It is a much larger protein with
influx and efflux have been studied in synaptosomes [14]
12 putative membrane spanning domains. It has an unu-
and hippocampal slices [7], but the mechanism of zinc
sually broad substrate range that includes Fe2+, Zn2+,
transport has never been elucidated. Studies with synapto-
Mn2+, Co2+, Cd2+, Cu2+, Ni2+, and Pb2+. The relationship
somes showed high affinity zinc uptake (K =
between recently-cloned mammalian zinc transporters and
uptake was independent of both ATP and the Na+ gradient
previously measured zinc fluxes is unclear at present.
[14]. Metal ion transporters have been studied in a variety of
The goal of the present studies was to provide much-
needed new information on the mechanism of zinc ioninflux and efflux across the plasma membrane of neurons.
* Tel.: +1 740 5930198; fax: +1 740 5930300;
Plasma membrane vesicles (PMV) from whole rat brain
1998 Elsevier Science Ireland Ltd. All rights reserved
R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
exchange in intact cells [8]. We have shown that in ourvesicle preparations, XIP was consistently able to inhibit
Na /Ca2+ exchange to 20% of control [15]. This findingindicates that our vesicle preparations are at least 80%inside-out.
The following studies were performed to distinguish vesi-
cular zinc uptake from zinc binding. After an uptake reac-tion was complete, the vesicles were washed three timeswith an ice-cold buffer containing 1 mM EGTA. This pro-cedure should remove most of the bound zinc but may notremove all of the bound zinc. To determine the amount ofbound zinc measured in our uptake reactions, zinc uptakewas measured under various conditions as described in Fig.
Fig. 1. Time-course of zinc uptake under various conditions. Plasma
1 and Table 1. Fig. 1 shows the time-course of zinc uptake
membrane vesicles were diluted into a buffer containing either 320
observed with 30 mM ZnCl2 added. Vesicles showed a rapid
mM sucrose (W), 137 mM choline (O), or 137 mM KCl (B) in 10 mMHEPES pH 7.4 at 37°C. Additional experiments included either 1 mM
accumulation of zinc during the first minute of a reaction.
La+ + + (A), or 1 mM O-phenanthroline (X) in the reaction buffer from
After 5 min vesicular zinc content began to plateau. Zinc
the start of the reaction. In the final experiment zinc uptake was
uptake was highly sensitive to temperature and was largely
measured in 137 mM KCl, 10 mM HEPES, pH 5.0 (L). Each point
represents the mean ± SE of three replicate determinations.
(27°C → 37°C) of 1.96 was calculated for the initial velo-city of zinc uptake, a value much greater than would be
were prepared as described previously [15] and stored in
expected for simple diffusion and binding reactions. Further
320 mM sucrose, 10 mM HEPES, pH 7.4 at −70°C until
confirmation that zinc uptake represented transported zinc is
use. Our procedure for subcellular fractionation results in
shown in Table 1 where it is shown that ionomycin (10 mM)
vesicular preparations that are highly enriched in plasma
inhibited zinc uptake. Addition of 1 mM O-phenanthroline
membranes. This conclusion is based on the assay of several
(a heavy metal chelator) to the binding reaction reduced
plasma membrane markers (e.g. [3H]-QNB binding (B
measurable zinc uptake almost to zero. La3+ (1 mM) and
0.6 pmol/mg), [3H]ouabain binding (B
H+ (buffer pH = 5.0) were shown to be effective inhibitors
without alamethicin addition), Na+/Ca2+ exchange transport
of zinc uptake but gave larger uptake values than that seen
with O-phenanthroline (see Fig. 1). The difference probably
uptake PMV were thawed on ice and diluted into either
represents zinc bound to the extravesicular surface. Zinc
320 mM sucrose or 137 mM KCl, 10 mM HEPES, pH 7.4
uptake was greatest in sucrose-containing media. Increasing
at 37°C containing various concentrations of ZnCl2 and
the ionic strength of the media with Na+, K+ or choline
zinc65 (3.2 mCi/ml). Uptake reactions were stopped by the
chloride reduced zinc uptake similarly (see Fig. 1, data for
addition of 1 mM LaCl3 and placing the reaction tubes on
Na not shown). This effect was not due to a competitive
ice. PMV suspensions are rapidly filtered on GF/C mem-
interaction between monovalent cations and zinc (see Fig.
branes and washed three times with an ice-cold solution of
2, below). Various transport inhibitors were ineffective
137 mM choline Cl/10 mM HEPES pH 7.4/1 mM EGTA.
at stopping zinc uptake (control zinc uptake (nmol/mg per
Zinc65 caught on the membranes was determined using a
30 s, mean ± SE): 6.99 ± 0.54; XIP (100 mM): 5.74 ± 0.51;
A determination of the relationship between zinc uptake
observed in these vesicle preparations and the influx and
Effect of valinomycin and ionomycin on zinc accumulation
efflux of zinc in the neuron is critical, but experimentallyproblematic. This is because vesicle preparations, although
providing an excellent system for kinetic studies, have a
mixture of inside-out and right side-out orientation. To
determine precisely the percentage of membrane vesicles
that are inside-out and right side-out is very difficult at
best. Fortunately, preparation techniques can sometimes
be devised that produce preparations that are of almostentirely one orientation. Exchange inhibitory peptide
Zinc binding was measured in 137 mM KCl/10 mM HEPES pH 7.4.
(XIP) is a highly selective inhibitor of Na /Ca2+ exchange
After 30 s the reaction was stopped by adding 1 mM La3+ and placing
in the brain [15]. XIP interacts with the cytoplasmic
the tubes on ice. Each compound was added to the reaction bufferbefore starting the reaction. Each value represents the mean ± SE
exposed portion of the protein. XIP is a highly-charged
obtained from at least three replicate experiments.
peptide and would not be expected to cross the plasma
*Significantly different when compared to control, one-way ANOVA
membrane and is totally ineffective at inhibiting Na+/Ca2+
(Tukey’s multiple comparison test), P Ͻ 0.05.
R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
amiloride (100 mM): 10.2 ± 0.06; DIDS (100 mM): 5.63 ±0.18). Interestingly, amiloride, an inhibitor of Na+/H+exchange, stimulated zinc uptake, albeit, this is likely tobe an indirect effect, owing to the high concentration (100mM) required.
Vesicular zinc uptake was saturable (see Fig. 2). Line-
weaver–Burk analysis of the initial velocity of zinc uptakeover a concentration range 1–300 mM yielded estimates forKm and Vmax of 14–16 mM and 10–12 nmol/mg per 30 s,respectively. Km and Vmax were not significantly affected if
K was substituted with Na2 + . Substitution with sucrose hadno effect on Km and only increased Vmax. Zinc uptake wasunaffected by preloading vesicles with Na+ or K+. Therefore,outwardly-directed gradients for these ions had no effect onzinc uptake (data not shown). Rapid uptake of zinc was seenin the absence of ATP. Valinomycin, a potassium-selectiveionophore, did not affect zinc uptake (see Table 1). Theresults are consistent with an electroneutral transport pro-cess, suggesting the transport of zinc may be coupled toother ions. The inhibition of zinc uptake seen when increas-
ing extravesicular H , is consistent with a Zn2+/2H+ antiportmechanism.
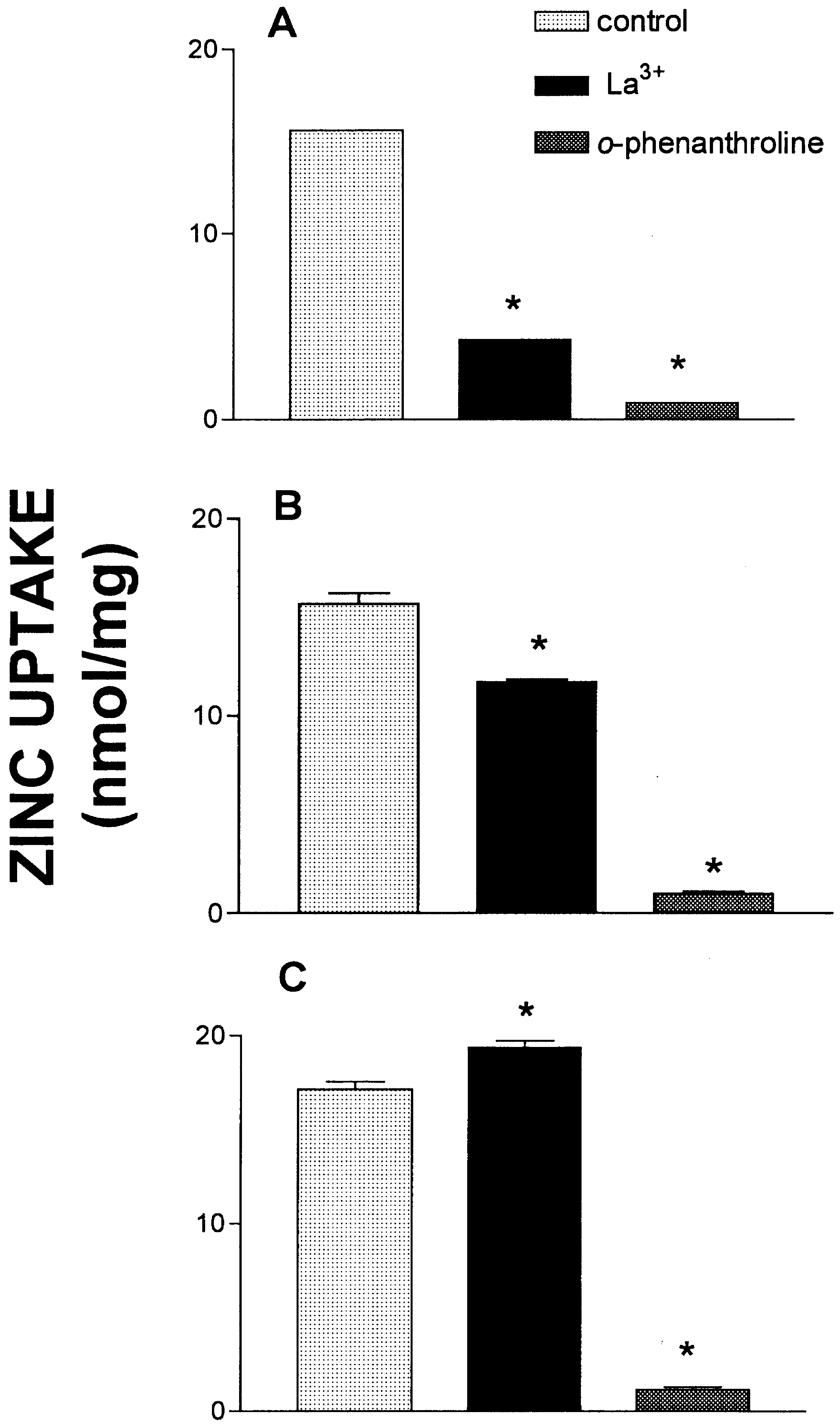
Fig. 3 shows data that suggests zinc uptake was indeed
reversible. In Fig. 3, zinc uptake was measured in KCl buf-
Fig. 3. Effect of La3+ and O-phenanthroline on zinc uptake. Zinc
fer. In panel A vesicles were exposed to each condition
uptake was measured in KCl buffer. (A) In these experiments each
(La3+, 1 mM or O-phenanthroline, 1 mM) from the start of
agent was added to the reaction mixture before the start of the
the reaction. As expected, zinc uptake was largely inhibited
reaction. Control indicates addition of 10 mM HEPES pH 7.4. The
in each condition, the difference between La3+ and O-phe-
reaction was allowed to proceed for 5 min, then the tubes were
nanthroline representing binding to the extravesicular sur-
placed on ice. (B) Each agent was added 30 s after the start of thereaction, then the reaction was stopped at 5 min by placing the tubes
face. When each condition was initiated 30 s after the start
on ice. (C) Each agent was added 5 min after the start of the reac-tion, then the tubes were allowed to incubate for an additional 5 minat 37°C before placing the tubes on ice. Each column represents themean ± SE of three replicate determinations. *, significantly differentwhen compared to control, one-way ANOVA (Tukey’s multiple com-parison test), P Ͻ 0.05.
of an uptake reaction (Fig. 3B), O-phenanthroline comple-tely reversed the zinc uptake that had occurred during thefirst 30 s of the reaction. This is shown by the differenceseen between La3+ and the O-phenanthroline. O-phenanthro-line, by lowering extravesicular zinc to near zero inducedthe rapid release of intravesicular zinc. La3+ was effective atstopping an ongoing uptake reaction, but this did not resultin a loss of accumulated zinc. In panel C each condition wasinitiated during the plateau phase, then each reaction wasallowed to proceed for an additional 5 min. Over the time-frame of the experiments (10 min at 37°C) the vesicles werestable and no evidence was obtained that the vesiclesbecame ‘leaky to zinc’ during an uptake reaction. Again,O-phenanthroline resulted in a complete loss of accumu-
Fig. 2. Lineweaver–Burk plot: zinc dependence of zinc uptake in
lated zinc. The release of accumulated zinc also was
various reaction buffers. Plasma membrane vesicles were diluted
induced by the addition of 1 mM EGTA (data not shown).
into a buffer containing either 137 mM KCl (L), 137 mM NaCl (A),
In contrast, La3+, which was effective at stopping uptake, did
or 320 mM sucrose (X) and 10 mM HEPES pH 7.4 at 37°C. The
initial velocity of zinc uptake was estimated by stopping the reaction
A complete understanding of the neuronal influx and
30 s after initiation with 1 mM La2++. The zinc concentrations usedwere 300 mM, 100 mM, 30 mM, 10 mM, 3 mM, and 1 mM.
efflux pathways for zinc is of critical importance before
R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
synaptic zinc homeostasis can be completely understood.
The technical assistance of Nancy Davis is gratefully
The cellular mechanisms of zinc efflux and influx are lar-
acknowledged. This work was supported by a grant from
gely unknown and few models exist that describe cellular
the Alzheimer’s Association and the American Heart
zinc transport. The findings of the present studies have iden-
Association, Ohio–West Virginia Affiliate.
tified a freely-reversible transport of zinc across the plasma
[1] Atwood, C.S., Moir, R.D., Huang, X., Tanzi, R.E. and Bush, A.I.,
membrane of neurons. Therefore, this zinc transporter could
Cerebral zinc metabolism in Alzheimer’s disease. In W. Wasco
mediate both zinc influx and zinc efflux in living cells. The
and R.E. Tanzi (Eds.), Molecular Mechanisms of Dementia,
zinc transporter mediates a rapid and large-capacity trans-
Humana Press, Totowa, NJ, 1997 pp. 225–237.
[2] Choi, D.W., Zinc neurotoxicity may contribute to selective neu-
levels are thought to be quite low (0.15 mM) [1] and peak
ronal death following transient global cerebral ischemia, ColdSpring Harb. Symp. Quant. Biol., 61 (1996) 385–387.
intracellular zinc concentrations have been recently esti-
[3] Cuajungco, M.P. and Lees, G.J., Zinc and Alzheimer’s disease:
mated in neurons to be between 35 and 45 nM [13]. How-
is there a direct link?, Brain Res. Rev., 23 (1997) 219–236.
ever, it is thought that concentrations as high as 300 mM are
[4] Eide, D., Molecular biology of iron and zinc uptake in
reached in the synaptic cleft after intense stimulation [2].
eukaryotes, Curr. Opin. Cell Biol., 9 (1997) 573–577.
Therefore, the zinc transporter may be involved in the reup-
[5] Erickson, J.C., Hollopeter, G., Thomas, S.A., Froelick, G.J. and
Palmiter, R.D., Disruption of the metallothionein-III gene in
take of zinc into the presynaptic fiber after stimulation to
mice: analysis of brain zinc, behavior and neuron vulnerability
prevent prolonged neurotoxic concentrations from occur-
to metals, aging, and seizures, J. Neurosci., 17 (1997) 1271–
ring extracellularly and to replenish synaptic vesicle stores.
Zinc influx would be driven by the large concentration gra-
[6] Gunshin, H., Mackenzie, B., Berger, U.V., Gunshin, Y.,
dient for zinc (micromolar extracellularly and nanomolar
Romero, M.F., Boron, W.F., Nussberger, S., Gollan, J.L. andHediger, M.A., Cloning and characterization of a mammalian
intracellularly). Between bursts of activity, the zinc trans-
proton coupled metal ion transporter, Nature, 388 (1997)
porter would be quiescent owing to its low affinity for zinc.
It is unlikely that the zinc transporter is an effective zinc
[7] Howell, G.A., Welch, M.G. and Frederickson, C.J., Stimulation
efflux protein, as this would require an affinity for zinc in
induced uptake and release of zinc in hippocampal slices,
the nanomolar range. Important to the overall goal of under-
[8] Li, Z.P., Nicoll, D.A., Collins, A., Hilgemann, D.W., Filoteo,
standing brain zinc homeostasis will be the identification of
a high affinity zinc transporter that can function in the 1–
Philipson, K.D., Identification of a peptide inhibitor of the car-
100 nM range of zinc concentrations. Metal ion transport
diac sarcolemmal Na+–Ca2 + Exchanger, J. Biol. Chem., 266
studies in yeast have consistently shown the existence of
high- and low-affinity transporters [4]. The recently-cloned
[9] Palmiter, R.D. and Findley, S.D., Cloning and functional char-
acterization of a mammalian zinc transporter that confers resis-
plasma membrane zinc efflux protein (ZnT-1) may mediate
tance to zinc, EMBO J., 14 (1995) 639–649.
high-affinity neuronal zinc efflux. The present findings
[10] Palmiter, R.D., Cole, T.B., Quaife, C.J. and Findley, S.D., ZNT-
demonstrate that the zinc transporter activity was markedly
3, a putative transporter of zinc into synaptic vesicles, Proc.
enhanced by lowering ionic strength. The ionic strength
Natl. Acad. Sci. USA, 93 (1996) 14934–14939.
effect is likely a reflection of the effect of membrane surface
[11] Reyes, J.G., Zinc transport in mammalian cells, Am. J. Physiol.,
charge on local ion concentration, as has been described for
[12] Richter, S., Hamann, J., Kummerow, D. and Bernhardt, I., The
the monovalent cation transporter in human erythrocytes
monovalent cation ‘leak’ transport in human erythrocytes: an
[12]. Finally, the present studies show that the zinc trans-
electroneutral exchange process, Biophys. J., 73 (1997) 733–
porter activity was inhibited by lowering extravesicular pH.
This finding rules out the possible involvement of DCT1
[13] Sensi, S.L., Canzoniero, L.M.T., Yu, S.P., Ying, H.S., Koh, J.-
Y., Kerchner, G.A. and Choi, D.W., Measurement of intracellu-
family members in mediating this zinc transport [6].
lar free zinc in living cortical neurons: routes of entry, J.
DCT1-mediated metal ion transport was H -coupled. In
contrast, the present findings show that zinc uptake was
blocked in the presence of micromolar concentrations of
Vandenhammer, C.J.A., Zinc uptake into synaptosomes, J.
H + . The zinc transporter activity is no doubt important in
[15] Wu, A. and Colvin, R.A., Characterization of exchanger inhibi-
maintaining neuronal zinc homeostasis and future studies
tory peptide (XIP) effects on Na+/Ca2+ exchange in rat and
will determine its involvement in the possible dysregulation
human brain plasma membrane vesicles., J. Neurochem., 63
of zinc levels in neurodegenerative diseases such as stroke,
severe epilepsy, and Alzheimer’s disease.
Sunglasses Safety FactSheetHS05-045A (9-05)Sunglasses aren’t just fashion accessories. They are a necessary protection for the eyes. Most consumers know about the danger of sun exposure to the skin, but many are unaware that the sun’s rays can damage the eyes. To cor-rectly shield the eyes, the right type of sunglasses must be worn, especially since wearing the wrong type can cause more
CENTRE FOR COMPANION ANIMAL HEALTH The School of Veterinary Science, The University of Queensland ww.uq.edu.au/ccah; +617 3365 2122 Dosing protocol for cats on glargine or detemir using daily home monitoring of blood glucose concentrations to adjust insulin dose Roomp K, RAND JS Evaluation of intensive blood glucose control using glargine in diabetic cats. Vet Intern Med 2

 Neuroscience Letters 247 (1998) 147–150
Characterization of a plasma membrane zinc transporter
Neuroscience Letters 247 (1998) 147–150
Characterization of a plasma membrane zinc transporter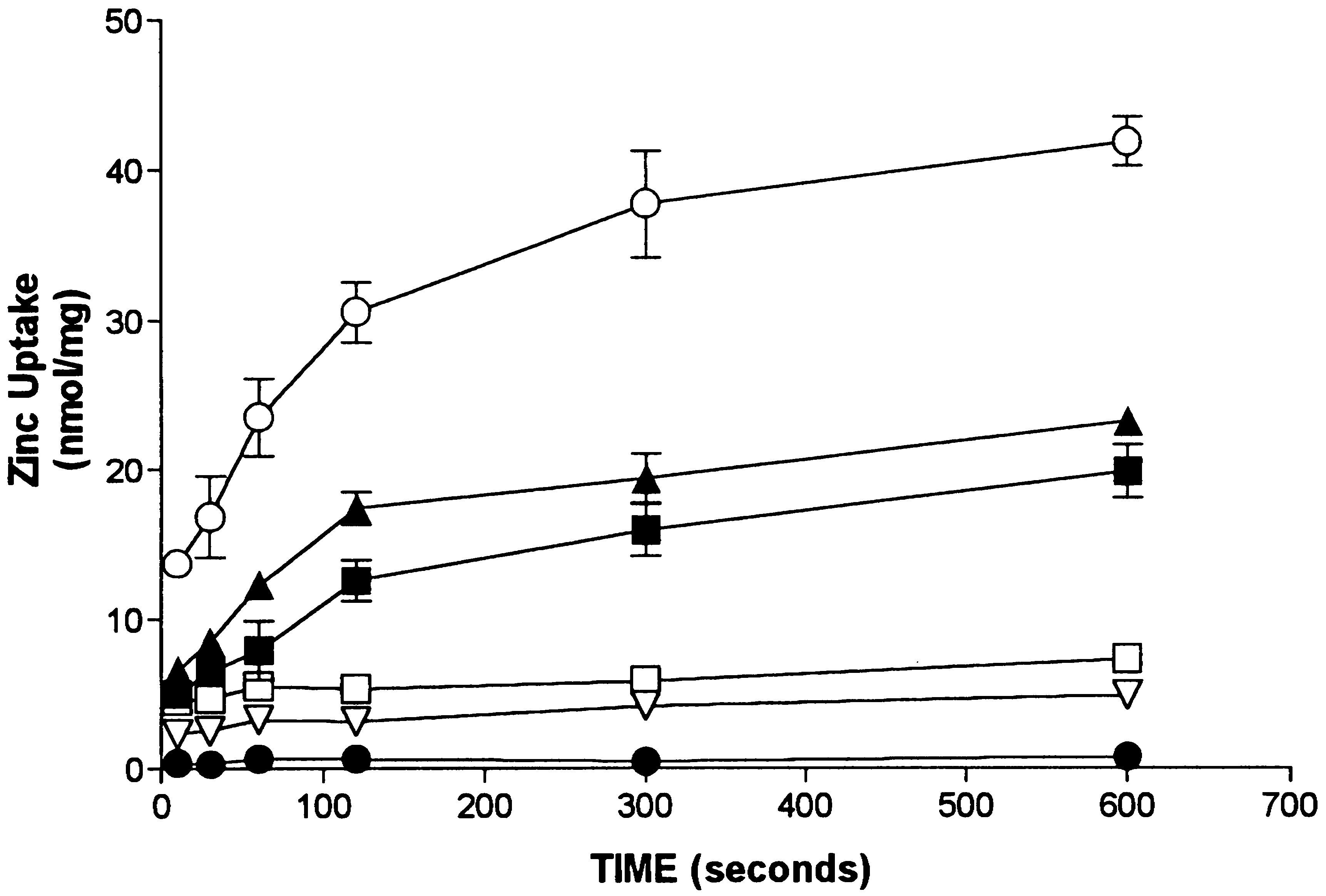 R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
exchange in intact cells [8]. We have shown that in ourvesicle preparations, XIP was consistently able to inhibit
Na /Ca2+ exchange to 20% of control [15]. This findingindicates that our vesicle preparations are at least 80%inside-out.
R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
exchange in intact cells [8]. We have shown that in ourvesicle preparations, XIP was consistently able to inhibit
Na /Ca2+ exchange to 20% of control [15]. This findingindicates that our vesicle preparations are at least 80%inside-out.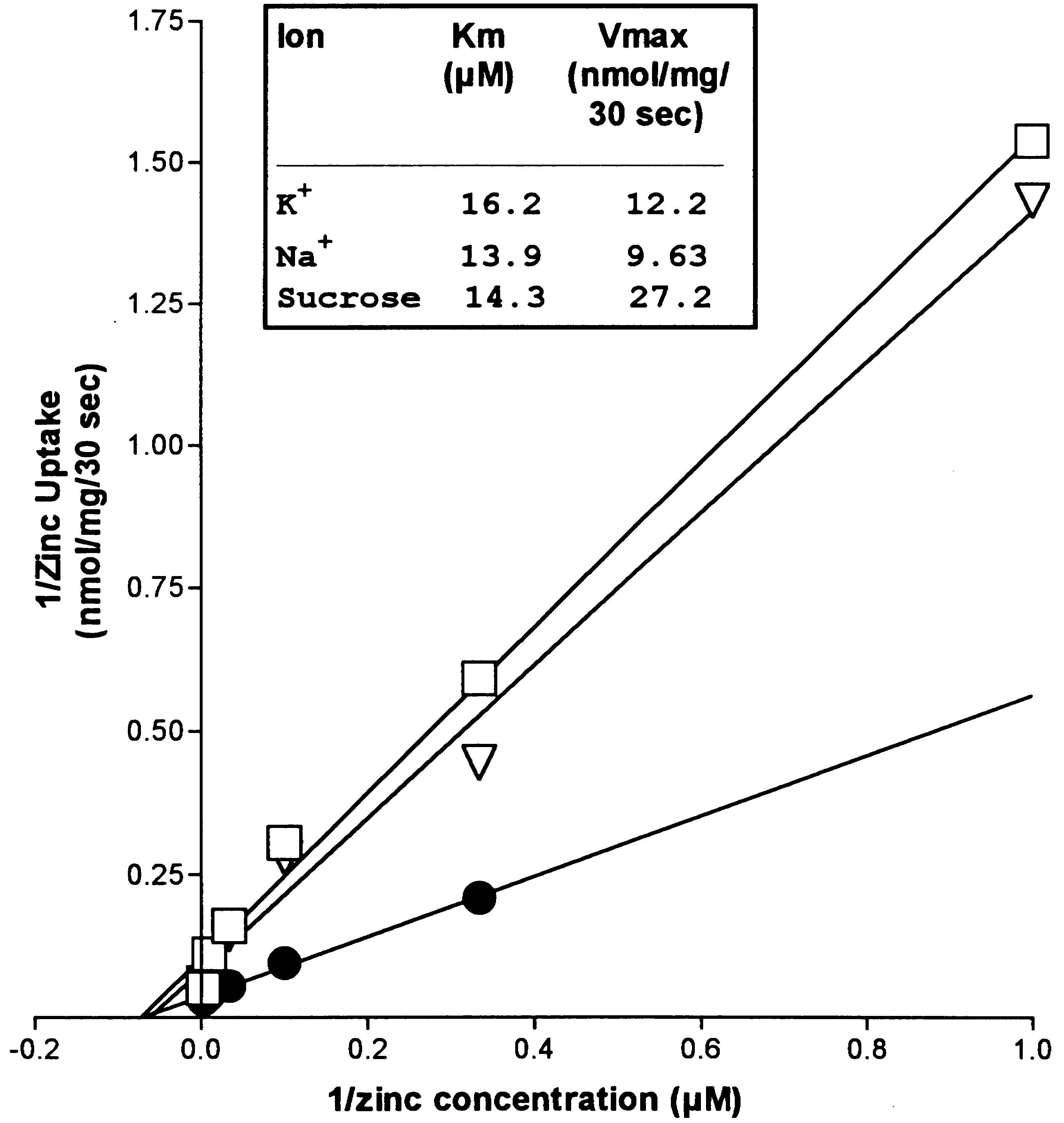
 R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
amiloride (100 mM): 10.2 ± 0.06; DIDS (100 mM): 5.63 ±0.18). Interestingly, amiloride, an inhibitor of Na+/H+exchange, stimulated zinc uptake, albeit, this is likely tobe an indirect effect, owing to the high concentration (100mM) required.
R.A. Colvin / Neuroscience Letters 247 (1998) 147–150
amiloride (100 mM): 10.2 ± 0.06; DIDS (100 mM): 5.63 ±0.18). Interestingly, amiloride, an inhibitor of Na+/H+exchange, stimulated zinc uptake, albeit, this is likely tobe an indirect effect, owing to the high concentration (100mM) required.