Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
No job name
Environ. Sci. Technol. 2001, 35, 2476-2481 Assessing the Biological Potency of
hormone biosynthesis, and/or by inducing the enzymesresponsible for steroid metabolism (1). Particular focus has
Binary Mixtures of Environmental
fallen on chemicals which mimic estradiol-17
binding to the estrogen receptor(s) (ERs) to elicit agonist
Estrogens using Vitellogenin
and/or antagonistic responses (2, 3). These chemicals are
Induction in Juvenile Rainbow Trout
collectively described as xenoestrogens. With the exceptionof the synthetic steroids, the xenoestrogens discovered so
(Oncorhynchus mykiss)
far are only weakly active when compared with endogenoussteroids. However, evidence for reproductive abnormalitiesthat are indicative of sex hormone disruption in wild fish
K A R E N L . T H O R P E , * , † , § T H O M A S H .
populations, supported by in situ monitoring studies with
H U T C H I N S O N , † M A L C O L M J .
caged fish, implies that some aquatic environments contain
H E T H E R I D G E , † M A R T I N S C H O L Z E , # J O H N
xenoestrogens at concentrations high enough to be of
P . S U M P T E R , ‡ A N D C H A R L E S R . T Y L E R §
concern to wildlife (4-6). These “field” observations are
Brixham Environmental Laboratory, AstraZeneca UK Limited,
supported by in vivo laboratory studies, where exposure to
Freshwater Quarry, Brixham, Devon, TQ5 8BA, U.K.,
some xenoestrogens induces estrogenic effects in fish at
School of Biological Sciences, The Hatherley Laboratories,
environmentally relevant concentrations (7-9). University of Exeter, The Prince of Wales Road, Exeter,
The aquatic environment receives a large influx of natural
Devon, EX4 4PS, U.K., Department of Biology and Chemistry,
and synthetic chemicals from agricultural, industrial, and
University of Bremen, Germany, and Fish Physiology Research
domestic sources. In Europe, natural estrogens such as E2
Group, Department of Biological Sciences, Brunel University,
and estrone, synthetic steroids including, 17R-ethynylestra-
Uxbridge, Middlesex, UB8 3PH, U.K.
diol, and other nonsteroidal chemicals known to haveestrogenic effects (such as alkylphenols), have been detectedin effluents that discharge into rivers (10, 11). This wide rangeof xenoestrogens in the aquatic environment highlights the
Experiments were conducted to assess the in vivo
importance of improving our understanding of combination
potency of binary mixtures of estrogenic chemicals using
effects of these chemicals in organisms as well as their
plasma vitellogenin (VTG) concentrations in juvenile
individual effects. In many cases xenoestrogens are present
rainbow trout (Oncorhynchus mykiss) as the endpoint.
at concentrations too low to be considered of concern
The estrogenic potencies of estradiol-17 (E2), 4-tert-
individually, but the presence of mixtures of these chemicals
nonylphenol (NP), and methoxychlor (MXC) were determined
means there is a potential for additive and/or interactive
following 14 day exposures to the individual chemicals
effects. The existence of interactive effects implies that the
and binary mixtures of these chemicals. E2, NP, and MXC
estrogenic effect of a mixture somehow deviates from whatis expected, on the basis of the estrogenic effects of the single
all induced concentration dependent increases in plasma
agents. There are two main analytical models for defining
VTG, with lowest observed effect concentrations of 4.7 and
the expected effects of a mixture: the model of concentration
7.9 ng L-1 for E2, 6.1 and 6.4 µg L-1 for NP, and 4.4 and
addition (CA), which assumes that the compounds act via
6.5 µg L-1 for MXC. Concentration-response curves for fixed
a similar mechanism in producing an effect (12), and the
ratio binary mixtures of E2 and NP (1:1000), E2 and MXC (1:
model of response addition, which assumes that the com-
1000), and NP and MXC (1:1) were compared to those
pounds act via independent pathways (13). If a mixture of
obtained for the individual chemicals, using the model of
xenoestrogens is more potent than would be expected, the
concentration addition. Mixtures of E2 and NP were additive
combination effect is described as more than additive
at the concentrations tested, but mixtures of E2 and
(synergistic), and if it is less effective, the combination effect
MXC were less than additive. This suggests that while NP
is described as less than additive (antagonistic). Suchdeviations from expectation should be demonstrated at more
probably acts via the same mechanism as E2 in inducing
than one concentration of the mixture.
VTG synthesis, MXC may be acting via a different mechanism-
In this study, the estrogenic activities of three environ-
(s), possibly as a result of its conversion to HPTE which
mental estrogens, namely E2, 4-tert-nonylphenol [NP], and
is an estrogen receptor R agonist and an estrogen receptor
methoxychlor [MXC], were assessed individually and in
antagonist. It was not possible to determine whether
binary mixtures (to investigate possible interactive effects)
mixtures of MXC and NP were additive using VTG induction,
using plasma vitellogenin (VTG) concentrations in juvenile
because the toxicity of MXC restricted the effect range
female rainbow trout (9), as the response. VTG induction in
for which the expected response curve for the binary mixture
fish is specifically an estrogen-dependent process, normally
could be calculated. The data presented illustrate that
restricted to mature females. During reproductive develop-
the model of concentration addition can accurately predict
ment in female fish, the hypothalamic-pituitary-gonadalaxis stimulates the ovary to produce E2, which is released
effects on VTG induction, where we know that both
into the bloodstream and transported to the liver. Here it
chemicals act via the same mechanism in mediating a
diffuses passively into hepatocytes and binds to the ER,
stimulating transcription of the VTG gene(s). The VTGsynthesized is then transported to the ovary and sequestered
Introduction A large number of natural and synthetic chemicals have been
* Corresponding author phone: (44) 1803-882882; fax: (44) 1803-
labeled as endocrine active, due to their ability to mimic
882974; e-mail: karen.thorpe@brixham.astrazeneca.com.
endogenous hormones. Endocrine active chemicals mediate
their effects by binding to hormone receptors as agonists or
antagonists, by inhibiting the enzymes responsible for steroid
2476 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 35, NO. 12, 2001
by the developing oocytes, to be stored as yolk for the
N-(tert-butyldimethylsilyl)-N-methylthrifluoroacetamide with
subsequent embryo (14). Although VTG synthesis is normally
1% tert-butyldimethylchlorosilane (MTBSTFA + 1% TBD-
restricted to maturing females, immature females and male
MCS). After cooling, 300 µL of bis(trimethysilyl)trifluoroac-
fish possess the machinery for VTG production, and exposure
etamide with 1% trimethylchlorosilane (BSTFA + 1% TMCS)
to estrogens (and their mimics) can trigger VTG synthesis via
was added, and the vial was heated to 60 °C for 20 min. The
the ER. Given that all the available evidence shows that steroid
reagents were removed under nitrogen, and the extracts were
estrogens and their mimics act via the ER to induce VTG
resuspended in 250 µL of dichloromethane. The derivatized
synthesis, the model of CA was used to assess the estrogenic
samples were analyzed on a Hewlett-Packard 6890 gas
activity of binary mixtures of the test chemicals. A simulation
chromatograph (GC) coupled to a Hewlett-Packard 5973 mass
technique termed “bootstrap” (15) was used with the model
spectrometer (MS), using helium as the carrier gas at 1 mL
of CA to construct a 95% confidence belt around the line of
min-1. The analysis conditions were as follows; sample
prediction. This provided a statistical basis to determine
volume, 1 µL; GC column, HP-5MS 30 m × 0.25 mm (id)
whether deviations from expectation were significant (those
fused silica with 0.25 µm film thickness; injector temperature,
that fall outside the 95% confidence of the predicted curve)
300 °C; column program, (1) 50 °C for 10 min, (2) increase
or simply due to natural variation in the biological response
to 300 °C at 8 °C min-1, (3) isothermal at 300 °C for 10 min.
(those that fall within the 95% confidence of the predicted
The MS was operated in the electron impact ionization mode
(70 eV) with selected ion monitoring (SIM) of the M-57 ionsfor the TBDMS/TMS derivative of E2 and its deuterated
Materials and Methods
analogue, ions m/z 401 and 403, respectively. Test Organisms. Female juvenile rainbow trout (approxi-
For measurement of NP, 1 L water samples were extracted
mately three months old) were obtained from West Country
under vacuum (50 mL min-1) onto preconditioned 47 mm
Trout, Trafalgar Farm, Cornwall, U.K. (experiments I and II)
C18 Empore disks (3M). NP was eluted from the disk using
and from Houghton Springs Fish Farm, Dorset, U.K. (experi-
10 mL of methanol, and the extract was diluted 1:1 with
ment III). The body weight of the fish used was 10.47 ( 0.71
HPLC grade water. Extracts were analyzed on a HPLC, using
g (mean ( SEM; n ) 24) in experiment I; 6.51 ( 0.48 g (n )
a Jasco PU980 LC pump with a mobile phase composition
24) in experiment II; and 7.89 ( 0.22 g (n ) 24) in experiment
of 80:20 methanol:water at 2 mL min-1. The analysis
III. In all experiments, fish were maintained for 14 days under
conditions were as follows; sample volume, 50 µL; HPLC
flow-through conditions in de-chlorinated water at 15.0 (
column, Hypersil H5ODS 150 mm × 4.6 mm (id) (Hichrom);
1 °C, with a 16 h light:8 h dark photoperiod, with 20 min
Jasco FP920 fluorescence spectrometer detection at 230 nm
dawn and dusk transition periods. Prior to the start of each
experiment, fish were acclimated in the same conditions for
For measurement of MXC, water samples were liquid/
a minimum of 10 days. Throughout the exposures, fish were
liquid extracted using hexane; extraction ratios for sample:
provided with a feeding ration of 1% of body weight per day
hexane were 80:1 (controls), 10:1 (2.4-7.5 µg L-1 treatments),
of Keystart Hatchery 1200 fish food pellets (BOCM Pauls
or 1:1 (13.5-24.0 µg L-1 treatment). Extracts were analyzed
on a Varian 3400 GC, using nitrogen as the carrier gas at 35
Test Chemicals. Methoxychlor (99% purity) was pur-
mL min-1. The analysis conditions were as follows: sample
chased from ChemService, Greyhound, Birkenhead, U.K. (Lot
volume, 1 µL; GC column, 1 m × 2 mm (i.d.) column packed
180-80A), technical grade NP (99% purity) was purchased
with 3% OV17 on 100/120 mesh (Phase Separations); injector
from Acros, Fisher Scientific, Loughborough, U.K. (Lot
temperature, 255 °C; column isothermal at 235 °C; electron
A010020701), and E2 (98% purity) was purchased from Sigma,
Experimental Design. Fish were exposed to five con- Water Supply and Test Apparatus. The supply of dechlo-
centrations of each of the individual chemicals and three
rinated water to the laboratory dosing system was monitored
concentrations of each binary mixture. Each experiment
daily for conductivity, hardness, and free chlorine and was
included a dilution water control (DWC) and a solvent control
tested for alkalinity and total ammonia twice weekly. The
(SC). In all experiments, each treatment consisted of a single
conductivity of the test water ranged from 204 to 238 µS
replicate containing 12 fish. The test vessels had a working
cm-1, the hardness from 41.3 to 47.7 mg L-1 (as CaCO
volume of 45 L and were constructed of glass, with a minimum
free chlorine was < 2 µg L-1. Alkalinity ranged from 21.2 to
of other materials (silicon rubber tubing and adhesive) in
Dissolved oxygen concentrations and pH levels were deter-
Stock solutions of each chemical were prepared weekly
mined in the individual tanks on days 0 and 1 and then twice
in HPLC grade methanol (Fisher Scientific) and dosed to
weekly throughout the exposure period. In all experiments,
glass mixing vessels by means of a peristaltic pump, at a rate
the dissolved oxygen concentration remained >80% of the
of 0.040 mL min-1, to mix with the dilution water flowing to
air saturation value throughout the exposures and pH values
the mixing vessels at a rate of 400 mL min-1. From each
ranged from 7.04 to 7.54. Water temperatures were monitored
primary mixing vessel the test solution flowed into a second
constantly throughout the exposure period and ranged from
mixing vessel to produce the binary mixtures and then into
15.2 to 15.8 °C. Dilution water and test chemical flow rates
the exposure tanks. The SC vessel received the same rate of
were checked at least three times per week. The flow-rate
addition of methanol, such that the water in all test vessels,
provided a 99% replacement time of approximately 7 h.
except the DWC, contained 0.01 mL methanol per liter. Analytical Chemistry. The actual concentrations of the
For each experiment a “fixed ratio” design was used for
reference chemicals were monitored throughout all experi-
the binary mixtures, in which the ratio of the two test
ments. Water samples were collected from each tank into
chemicals was kept constant, while the total concentration
solvent-cleaned flasks on days 0, 7, and 14 of the exposures.
of the mixture was varied. The ratio used for each experiment
For measurement of E2, 2.5 L water samples were spiked
was selected on the basis of earlier work in which E2 was
with 5 ng L-1 deuterated-E2 and then extracted under vacuum
found to be approximately 1000-fold more potent than NP
(50 mL min-1) onto preconditioned 47 mm C18 Envi-disks
(Supelco). E2 was eluted from the disks using 30 mL of
Experiment I - E2 + NP. Groups of 12 juvenile female
methanol, and the residual solvent was removed under a
rainbow trout were exposed for 14 days to nominal con-
stream of nitrogen. The extracts were derivatized by heating
centrations of E2 at 2.4, 4.2, 7.5, 13.5, and 24.0 ng L-1 and NP
to 120 °C for 20 min with 200 µL of pyridine and 300 µL of
at 2.4, 4.2, 7.5, 13.5, and 24.0 µg L-1 and to binary mixtures
VOL. 35, NO. 12, 2001 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2477 TABLE 1. Mean Measured Tank Concentrations of Test Chemicals, in the Individual and Binary Mixture Exposures over the 14 Daysa Experiment I. 17 -Estradiol and 4-tert-Nonylphenol mean measured concn estradiol (ng/L) mean measured concn nonylphenol (µg/L) individual individual Experiment II. 17 -Estradiol and Methoxychlor mean measured concn estradiol (ng/L) mean measured concn methoxychlor (µg/L) individual individual Experiment III. 4-tert-Nonylphenol and Methoxychlor mean measured concn nonylphenol (µg/L) mean measured concn methoxychlor (µg/L) individual individual
of E2 + NP at concentrations of 4.2 ng L-1 + 4.2 µg L-1, 7.5
The model parameter θmin describes the minimal mean
ng L-1 + 7.5 µg L-1, and 13.5 ng L-1 + 13.5 µg L-1, respectively.
effect (control response), θmax is the asymptotical maximal
Experiment II - E2 + MXC. Groups of 12 juvenile female
effect, θ1 is termed the “location” parameter, and θ2
rainbow trout were exposed for 14 days to nominal con-
characterizes the “steepness” of the concentration response
centrations of E2 at 2.4, 4.2, 7.5, 13.5, and 24.0 ng L-1 and
relationship. The experiments were not designed to deter-
MXC at 2.4, 4.2, 7.5, 13.5, and 24.0 µg L-1 and to binary
mine maximal effects, so estimation of θmax contains a high
mixtures of E2 + MXC at concentrations of 4.2 ng L-1 + 4.2
degree of statistical uncertainty. Due to heterogeneous
µg L-1, 7.5 ng L-1 + 7.5 µg L-1, and 13.5 ng L-1 + 13.5 µg L-1,
nonrandom variabilities in the replicated data (heterosce-
dasticity), each model was fitted using the estimation method
Experiment III - MXC + NP. Groups of 12 juvenile female
of generalized least squares (15). To fulfill the statistical
rainbow trout were exposed for 14 days to nominal con-
prerequisite of symmetrically distributed effect data for this
centrations of MXC at 2.4, 4.2, 7.5, 13.5, and 24.0 µg L-1 and
estimation method, the plasma VTG concentrations were
4-NP at 2.4, 4.2, 7.5, 13.5, and 24.0 µg L-1 and to binary
log10-transformed. LOECs were determined using a non-
mixtures of MXC + NP at concentrations of 4.2 µg L-1 + 4.2
parametric Wilcoxon’s rank sum test (17). µg L-1, 7.5 µg L-1 + 7.5 µg L-1, and 13.5 µg L-1 + 13.5 µg L-1,
The expected concentration-response relationships of
the binary mixtures were determined using the model of CA
Fish Sampling. In all experiments, a subgroup of fish (n
(12). The model can only be used to calculate mixture effects
) 24) was sampled at the outset (day 0) of the experiment,
for the same effect range observed for the individual
and then all exposed fish were sampled on day 14. Fish were
components of the mixture, and if these effects are quan-
sacrificed in a lethal dose (200 mg L-1) of MS222 (3-
titatively describable in a statistically valid way. Therefore,
aminobenzoic acid ethyl ester, methanesulfonate salt) (Sigma),
the expected curve of the binary mixtures can only be
buffered with 1 M NaOH to pH 7.3. Blood was collected by
calculated up to an effect range which is determined by the
cardiac puncture, using a heparinized syringe (5000 Units
minimum of the two estimated model parameter θmax of both
heparin mL-1) and centrifuged (7000g; 5 min, 15 °C), and the
compounds. The concentration-response curve for VTG
plasma was removed and stored at -80 °C until required for
induction is very steep, covering several orders of magnitude
VTG analysis. Plasma samples were assayed for VTG using
(ng/mL to mg/mL) and often results in a high variability in
an established homologous rainbow trout RIA (16).
the vitellogenic response between individuals in a treatment. Statistical Analyses. For the description of the concen-
Such an inherent variability can complicate any analysis on
tration effect relationships for the individual test compounds
exposure to individual or mixtures of chemicals. For this
and for the binary mixtures, a four-parameter logit regression
reason the “bootstrap” methodology (15) was employed with
the model of CA, to determine the statistical accuracy of thepredicted combined effects. Results and Discussion Measured Concentrations of the Test Chemicals. Mean
where x ) concentration and f(x) ) mean effect.
measured tank concentrations of the individual chemicals
2478 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 35, NO. 12, 2001 FIGURE 1. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles) and 4-tert-nonylphenol (NP) (open circles) and (b) fixed ratio binary mixtures (1:1000) of E2 and NP (open circles). In some cases the plasma VTG concentrations were very similar between fish within a treatment; therefore, not all data points are visible. For each of these exposures the 95% confidence belts (light gray shaded regions) of the fitted concentration-response relationships (gray lines) are shown. The expected vitellogenic response for the binary mixture, calculated using the model of concentration addition, is shown as a black line (b) with a 95% confidence bootstrap belt (dark gray shaded region).
and their binary mixtures are given in Table 1. All exposure
estrogen and as an antiandrogen in male fathead minnows
concentrations are described using means of the actual
(18) and in recombinant yeast reporter gene assays (3), and
measured concentrations. For most chemical exposures the
it also affects some of the cytochrome p450 mixed function
mean measured concentrations were between 70 and 110%
oxidase enzymes (19). Some estrogenic activity of NP may
of nominal. However, for the highest concentration of NP,
also result from alterations it can cause in concentrations of
24 µg L-1, the mean measured concentration was between
endogenous E2, rather than by effects mediated via the ER
51% and 68% of nominal. In experiment II (E2 + MXC), the
directly (20). The concentration additive behavior of NP with
mean measured concentration of E2 was 190% of nominal
E2 in these experiments, however, together with the ability
for the lowest test concentration (2.4 ng L-1), and the mean
of NP to bind the ER (but not the AR) in rainbow trout liver
measured concentrations of MXC were between 60% and
and brain cells (21), suggests that the VTG response induced
by NP is mediated via the ER alone. This may also be true
Plasma Concentrations of VTG in Control Fish. The
for the related alkylphenolic chemical, octylphenol, which
concentrations of VTG in the plasma of juvenile female fish
has been shown to act in an additive manner with E2, in
at the onset of the experiments were 500 ( 80 ng mL-1, 310
inducing VTG synthesis in rainbow trout (22).
( 50 ng mL-1, and 540 ( 80 ng mL-1 for experiments I, II,
Mixtures of E2 + MXC. For fish exposed to 13.0 µg L-1
and III, respectively. There were no detectable increases in
MXC there was a 58% mortality by day 14, and for fish exposed
plasma VTG concentrations in either the DWC or in the SC
to the mixture of 13.2 µg L-1 MXC + 12.8 ng L-1 E2 there was
fish after the 14 day exposure period in any of the experiments
a 25% mortality. In all other chemical exposures, 100% of the
fish were alive at the end of the experiment. A previous study
Mixtures of E2 and NP. There were no mortalities in fish
reported a 96 h median lethal concentration for MXC of 31.2
exposed to E2 and NP or in fish exposed to binary mixtures
µg L-1 in juvenile rainbow trout (23). The toxicity of MXC has
of these chemicals. Estradiol-17 (concentrations ranging
been demonstrated to increase with duration of exposure,
from 2.3 to 21.3 ng L-1) and NP (concentrations ranging from
and this may account for the apparent greater toxicity of
1.8 to 12.2 µg L-1) produced concentration-dependent
MXC observed in this study, compared with Heming et al.
increases in plasma VTG (Figure 1), with LOECs of 4.7 ng L-1
(plasma VTG concentration of 4100 ( 940 ng mL-1, p < 0.05)
In experiment II, E2 (concentrations ranging from 4.2 to
and 6.1 µg L-1 (plasma VTG concentration of 1450 ( 230 ng
23.0 ng L-1) and MXC (concentrations ranging from 1.4 to
mL-1, p < 0.05) for E2 and NP, respectively. The mixture of
13.0 µg L-1) produced concentration-dependent increases
E2 and NP, at a fixed 1:1000 ratio, also produced a
in plasma VTG (Figure 2), with LOECs of 7.9 ng L-1 for E2
concentration-dependent increase in plasma VTG, with the
(plasma VTG concentration of 22 460 ( 8660 ng mL-1, p <
lowest mixture concentration tested (4.9 ng L-1 E2 and 3.3
0.05) and 4.4 µg L-1 for MXC (plasma VTG concentration of
µg L-1 NP) inducing a 17-fold increase in VTG concentration
9340 ( 6590 ng mL-1, p < 0.05). The mixture of E2 and MXC,
(8720 ( 4370 ng mL-1, p < 0.05). The observed VTG induction
at a fixed 1:1000 ratio, also produced a concentration-
data for the mixture of E2 and NP was close to that predicted
dependent increase in plasma VTG (Figure 2), with the lowest
by the model of CA (Figure 1). The 95% confidence belt of
mixture concentration tested (4.3 ng L-1 E2 and 3.8 µg L-1
the fitted concentration-response relationship for the
MXC) inducing a plasma VTG concentration of 18340 ( 7680
observed mixture data overlapped with the 95% confidence
ng mL-1 (p < 0.05). When comparing the measured VTG
bootstrap belt for the calculated mean of CA, for the whole
induction that occurred for the mixture, with the expected
mixture effects, calculated according to the model of
For many estrogen mimics the pathways by which they
concentration addition, mixtures of E2 and MXC were shown
alter estrogen-sensitive pathways have yet to be fully
to act in a less than additive manner (Figure 2). The
characterized. NP has been shown to have multiple mech-
concentration-response curve for the experimental mixture
anisms of action on the endocrine system; it acts as both an
of E2 and MXC is displaced to the right of the predicted
VOL. 35, NO. 12, 2001 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2479 FIGURE 2. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles) and methoxychlor (MXC) (open circles) and (b) fixed ratio binary mixtures (1:1000) of E2 and MXC (open circles). In some cases the VTG concentrations were very similar between fish within a treatment; therefore, not all data points are visible. For each of these exposures the 95% confidence belts (light gray shaded regions) of the fitted concentration-response relationships (gray lines) are shown. The expected vitellogenic response for the binary mixture, calculated using the model of concentration addition, is shown as a black line (b) with a 95% confidence bootstrap belt (dark gray shaded region). FIGURE 3. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) methoxychlor (MXC) (closed circles) and 4-tert-nonylphenol (NP) (open circles) and (b) fixed ratio binary mixtures (1:1) of MXC and NP (open circles). In some cases the VTG concentrations were very similar between fish within a treatment; therefore, not all data points are visible. For each of these exposures the 95% confidence belts (light gray shaded regions) of the fitted concentration-response relationships (gray lines) are shown. The expected vitellogenic response for the binary mixture, calculated using the model of concentration addition, is shown as a black line (b) with a 95% confidence bootstrap belt (dark gray shaded region).
concentration-response curve, and there is no overlap
Although, ER has not yet been identified in the rainbow
between the two 95% confidence belts for the whole effect
trout, it has been identified in another species of teleost, the
seabream (26). In the seabream, ERR and ER are coexpressed
The model of CA is based on the assumption that the
in the liver, although whether they are both involved in the
components of a mixture mediate their effects by similar
regulation of vitellogenesis has still to be investigated. If,
modes of action. The small but significant deviation from
however, both ERR and ER are present in the liver of rainbow
additivity for the mixture of E2 and MXC suggests that MXC
trout and both play a role in the regulation of vitellogenesis,
mediated a vitellogenic response via a different mechanism
then the mixed agonist/antagonist activity of HPTE, in the
(or pathway) to E2. This complies with reports in the literature
presence of E2, might well account for the observed deviation
that MXC does not interact directly with the ER, in the same
way as E2. The estrogenic activity of MXC is thought to result
Mixtures of NP + MXC. There was 100% mortality of fish
from its primary metabolite, 2,2-bis-(p-hydroxyphenyl)-1,1,1-
exposed to MXC at a concentration of 20.2 µg L-1 after 6 days
trichloroethane (HPTE), which is produced in the liver (by
and 50% mortality in fish exposed to 14.3 µg MXC L-1 after
O-demethylation) (24). In mammals, HPTE has been found
14 days. In fish exposed to the binary mixture of 11.4 µg L-1
to act as an agonist for ERR but as an antagonist for ER (25).
MXC + 11.9 µg L-1 NP, there was a 58% mortality after 14
2480 9 ENVIRONMENTAL SCIENCE & TECHNOLOGY / VOL. 35, NO. 12, 2001
days, but in all other chemical exposures all of the fish were
even simple mixtures of environmental estrogens, on a clearly
defined endpoint with a known mechanism of action.
NP (concentrations ranging from 1.8 to 16.3 µg L-1) and
MXC (concentrations ranging from 2.0 to 20.2 µg L-1)
Acknowledgments
produced concentration-dependent increases in plasma VTG
This research was co-funded by the UK Environment Agency,
(Figure 3), with LOECs of 6.4 µg L-1 for NP (plasma VTG
National Centre for Ecotoxicology and Hazardous Substances,
concentration of 128 760 ( 117 540 ng mL-1, p < 0.05) and
Wallingford and the AstraZeneca Shared Research Program
6.5 µg L-1 for MXC (plasma VTG concentration of 16 390 (
(supported by AstraZeneca, Avecia, ICI and ZENECA Agro-
7160 ng mL-1, p < 0.05). The mixture of NP and MXC, at a
chemicals), Brixham Environmental Laboratory, Devon. The
fixed 1:1 ratio, produced a concentration-dependent increase
authors wish to acknowledge the valuable technical assistance
in plasma VTG, with the lowest mixture concentration tested
of colleagues at Brixham Environmental Laboratory, espe-
(3.0 µg L-1 NP and 3.5 µg L-1 MXC) inducing a plasma VTG
cially, Rob Cumming, Nadine Pounds, and Colin Woods.
concentration of 723 055 ( 481 814 ng mL-1 (p < 0.05). Determination of the predicted effects of the binary mixtureon the basis of concentration addition depends on the
Literature Cited
concentration-response curves derived for the individual
(1) LeBlanc, G. A.; Bain, L. J.; Wilson, V. S. Mol. Cell. Endocrinol.
compounds. Due to the toxicity of MXC in our study, only
2000, 126, 1.
a partial concentration-response curve was obtained, re-
(2) Danzo, B. J. Environ. Health Persp. 1997, 105, 294. (3) Sohoni, P.; Sumpter, J. P. J. Endocrin. 1998, 158, 327.
stricting the effect range for which the expected response
(4) Harries, J. E.; Sheahan, D. A.; Jobling, S.; Matthiessen, P.; Neall,
curve for the binary mixture could be calculated (Figure 3).
P.; Routledge, E. J.; Rycroft, R.; Sumpter, J. P.; Tylor, T. Environ.
At the lowest mixture concentration tested (4.2 µg L-1 NP +
Toxicol. Chem. 1996, 15, 1993.
4.2 µg L-1 MXC), the 95% confidence belt of the fitted
(5) Harries, J. E.; Sheahan, D. A.; Jobling, S.; Matthiessen, P.; Neall,
concentration-response relationship for the observed mixed
P.; Sumpter, J. P.; Tylor, T.; Zaman, N. Environ. Toxicol. Chem.
data overlapped with the 95% confidence bootstrap belt for
1997, 16, 534.
the calculated mean of CA, suggesting that NP and MXC
(6) Jobling, S.; Nolan, M.; Tyler, C. R.Brighty, G.; Sumpter, J. P. Environ. Sci. Technol. 1998, 32, 2498.
were acting in an additive manner. However, it was not
(7) Jobling, S.; Sheahan, D.; Osborne, J. A.; Matthiessen, P.; Sumpter,
possible to confirm or negate this for mixtures of NP and
J. P. Environ. Toxicol. Chem. 1996, 15, 194.
MXC at the higher concentrations tested (because of fish
(8) Panter, G. H.; Thompson, R. S.; Sumpter, J. P. Aquat. Toxicol.
mortality). On the basis of the findings for the mixture of E2
1998, 42, 243.
and MXC, the mixture of NP and MXC, would be expected
(9) Thorpe, K. L.; Hutchinson, T. H.; Hetheridge, M. J.; Sumpter,
to be less than additive. Having said this, the effects of NP
J. P.; Tyler, C. R. Environ. Toxicol. Chem. 2000, 19, 2812.
on cytochrome P450 mixed function oxidase enzymes (19)
(10) Desbrow, C.; Routledge, E. J.; Brighty, G.; Sumpter, J. P.; Waldock,
M. Environ. Sci. Technol. 1998, 32, 1549.
might affect the conversion of MXC to HPTE and so affect
(11) Blackburn, M. A.; Waldock, M. J. Water Res. 1995, 29, 1623.
the estrogenic activity of the mixture of NP and MXC.
(12) Loewe, S.; Muischnek, H. Arch. Exp. Pathol. Pharmakol. 1926,
In each of the experiments, VTG induction occurred at
the lowest mixtures concentrations tested, even when the
(13) Bliss, C. I. Ann. J. Appl. Biol. 1939, 26, 585.
components of the mixtures were present at concentrations
(14) Tyler, C. R. In Reproductive Physiology of fish; Scott, A. P.,
Sumpter, J. P., Kime, D. A., Rolfe, M. S., Eds.; Fish Symp. 91:
below their individual LOECs. These binary mixture con-
centrations, at which effects on plasma VTG occurred, reflect
(15) Scholze, M.; Boedeker, W.; Faust, M.; Backhaus, T.; Altenburger,
environmentally relevant concentrations of the chemicals
R.; Grimme, L. H. Environ. Toxicol. Chem. 2001, 20, 448.
in some aquatic environments. Surveys of domestic effluents
(16) Sumpter, J. P. In Current Trends in Comparitive Endocrinology;
in the UK have detected concentrations of E2 ranging from
Lofts, B., Holmes, W. M., Eds.; Hong Kong University Press:
1 to 88 ng L-1 (10, 27, 28) and concentrations of NP ranging
from 1 to 5 µg L-1 (11, 28). These effluents discharge directly
(17) Sokal, R. R.; Rohlf, F. J. Biometry; W. H. Freeman and Company:
into rivers, where measured concentrations range from 1 to
(18) Harries, J. E.; Runnalls, T.; Hill, E.; Harris, C.; Maddix, S.; Sumpter,
5 ng L-1 for E2 (11) and from 1 to 2.3 µg L-1 for NP (11). MXC
J. P.; Tyler, C. R. Environ. Sci. Technol. 2000, 34, 3003.
is no longer used in Europe, but it is still used extensively in
(19) Lee, P. C.; Patra, S. C.; Stelloh, C. T.; Lee, W.; Struve, M. Biochem.
the USA and Canada as well as in many third world countries,
Pharmacol. 1996, 52, 885.
to control biting fly larvae. Due to the large number of these
(20) Giesy, J. P.; Pierens, S. L.; Snyder, E. M.; Miles-Richardson, S.;
and other oestrogenic chemicals present in the aquatic
Kramer, V. J.; Snyder, S. A.; Nichols, K. M.; Villeneuve, D. A.
environment, additivity at concentrations below the LOECs
Environ. Toxicol. Chem. 2000, 19, 1368.
(21) Knudsen, F. R.; Pottinger, T. G. Aquatic Toxicol. 1999, 44, 159.
for the individual chemicals needs to be considered when
(22) Knudsen, F. R.; Arukwe, A.; Pottinger, T. G. Environ. Pollut.
assessing the possible implications of endocrine active
1998, 103, 75.
chemicals in the aquatic environment. The estrogenic
(23) Heming, T. A.; Sharma, A.; Kumar, Y. Environ. Toxicol. Chem.
potency of mixtures of chemicals discharged into the
1989, 8, 923.
environment is, therefore, important in any risk assessment
(24) Bulger, W. H.; Muccitelli, R. M.; Kupper, D. Biochem Pharmacol.
strategy. Although the applicability of the concept of CA has
1978, 27, 2417.
so far not been applied to the field of endocrine disruption,
(25) Gaido, K. W.; Leonard, L. S.; Maness, S. C.; Hall, J. M.; McDonnell,
D. P.; Saville, B.; Safe, S. Endocrinol. 1999, 140, 5746.
the data from these experiments suggest that it is suitable
(26) Socorro, S.; Power, D. M.; Olsson, P.-E.; Canario, A. V. M. J.
for the prediction of combination effects, where the com-
Endocrinol. 2000, 166, 293.
ponents of the mixture act through the same mechanism of
(27) Belfroid, A. C.; Van der Horst, A.; Vethaak, A. D.; Schafer, A. J.;
action on a clearly defined endpoint. The model accurately
Rijs, G. B. J.; Wegener, J.; Cofino, W. P. Sci. Total Environ. 1999,
predicted the combination effects on VTG induction for the
mixture of E2 and NP, where we know that both chemicals
(28) Rodgers-Gray, T. P.; Jobling, S.; Morris, S.; Kelly, C.; Kirby, S.;
Janbakhsh, A.; Harries, J. E.; Waldock, M. J.; Sumpter, J. P.; Tyler,
act via the ER in mediating a vitellogenic response. For the
C. R. Environ. Sci. Technol. 2000, 34, 1521.
mixture of E2 and MXC the model of CA overpredicted thecombination effect, but this is perhaps not surprising given
Received for review October 13, 2000. Revised manuscript
the complex mechanism by which MXC has been reported
received March 23, 2001. Accepted March 25, 2001.
to elicit an estrogenic response (24, 25). Our studies furtherhighlight the difficulties in assessing combination effects of
VOL. 35, NO. 12, 2001 / ENVIRONMENTAL SCIENCE & TECHNOLOGY 9 2481
CASE STUDY #4 Performance of Masimo SET® Pulse Oximetry in a Child with Meningococcemia A 2 month old male with meningococcemia was admitted to the pediatric intensive care unit (PICU) of a 242 bed regional medical center. Following admission, he developedrespiratory failure, septic shock and DIC (disseminated intravascular coagulation), whichprogressed to renal failure. He required hem
205 S. Whiting Street, Suite 303 703-823-1533 Solo private gynecology practice in Alexandria, Virginia since 1988, with emphasis on hormonal issues and preventive medicine. OB-GYN residency at the Washington Hospital Center, Wash. DC, 1979-1983. MD degree from University of Maryland School of Medicine, 1979. BS degree in Psychology from Duke University, 1975. Certified Menopause Practi
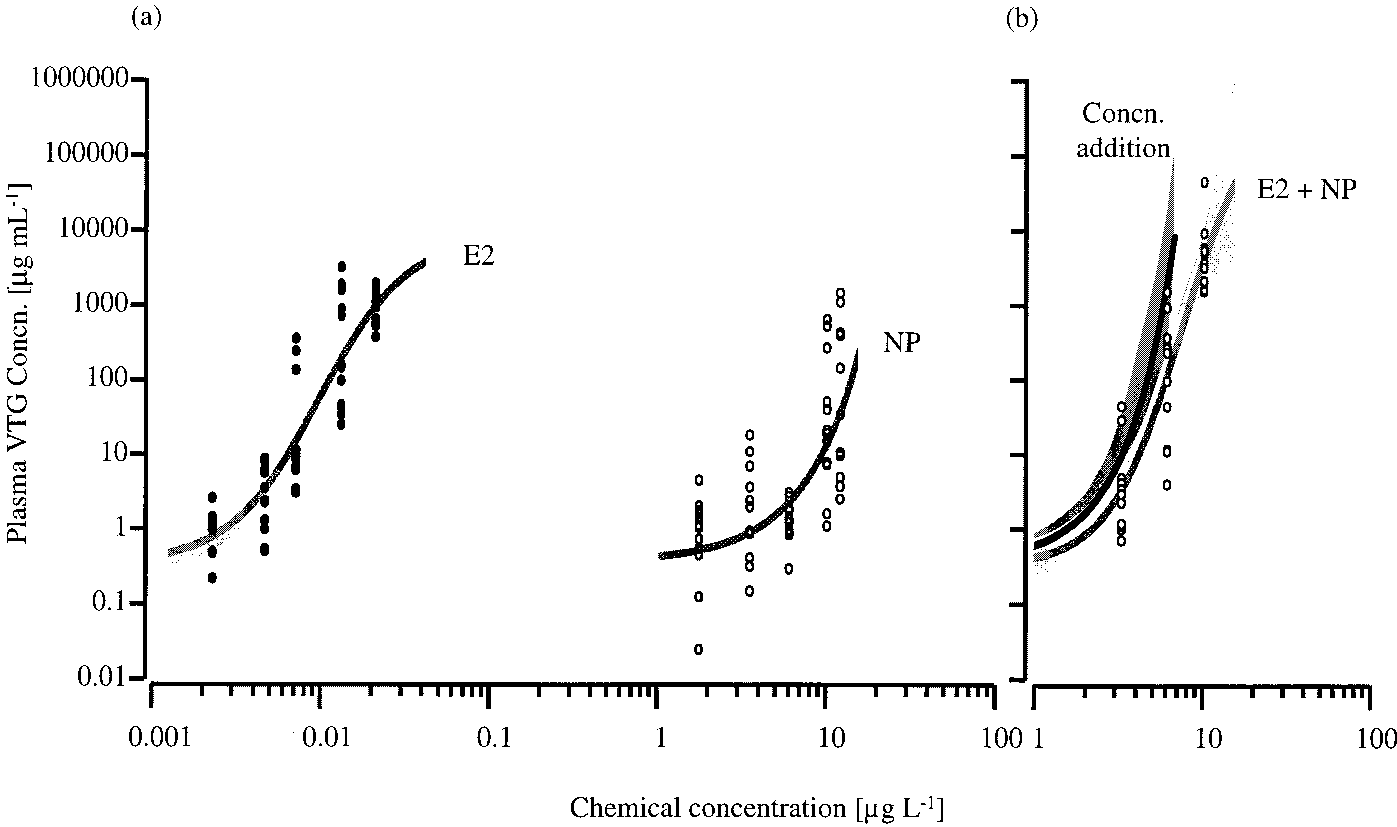 FIGURE 1. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles)
FIGURE 1. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles)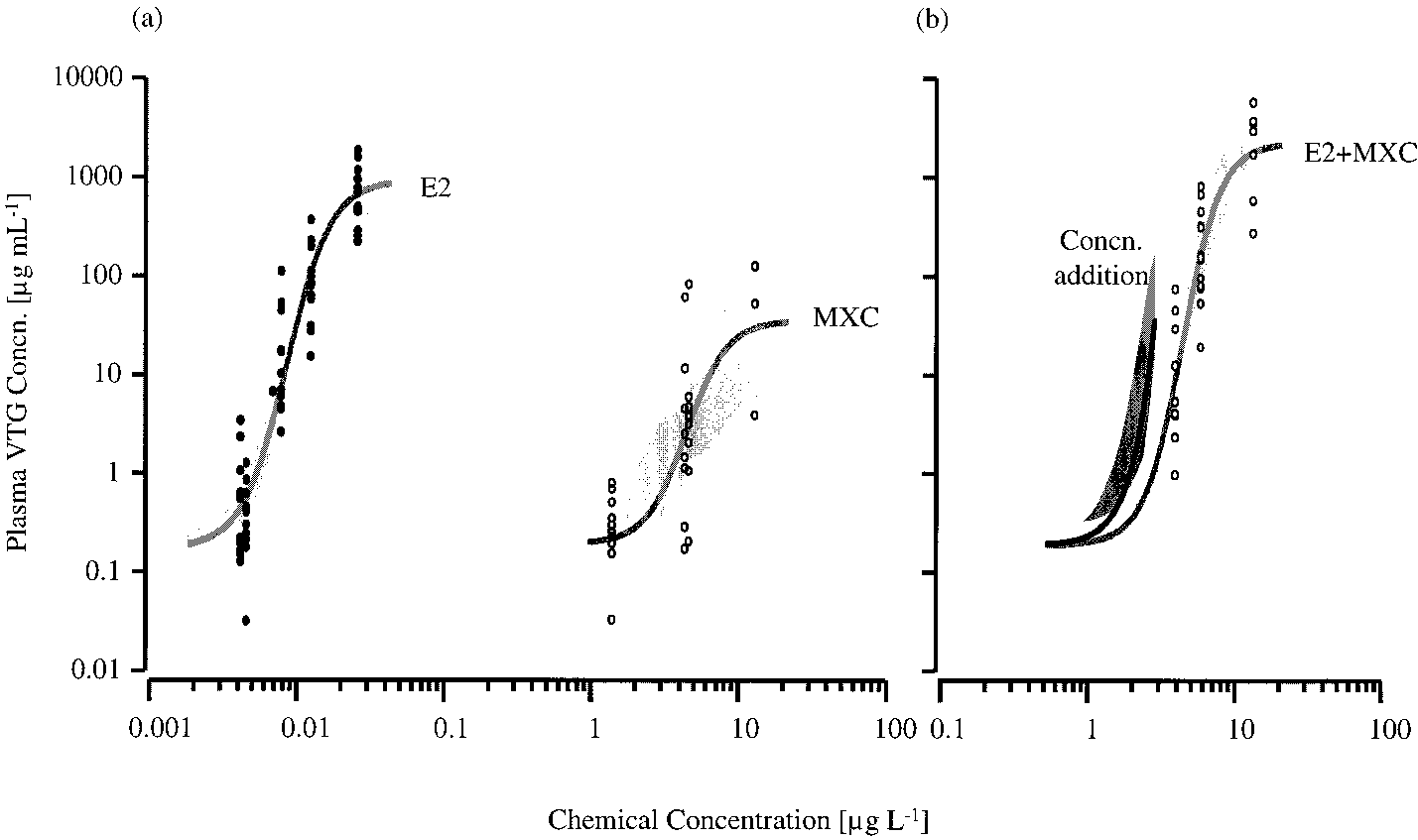
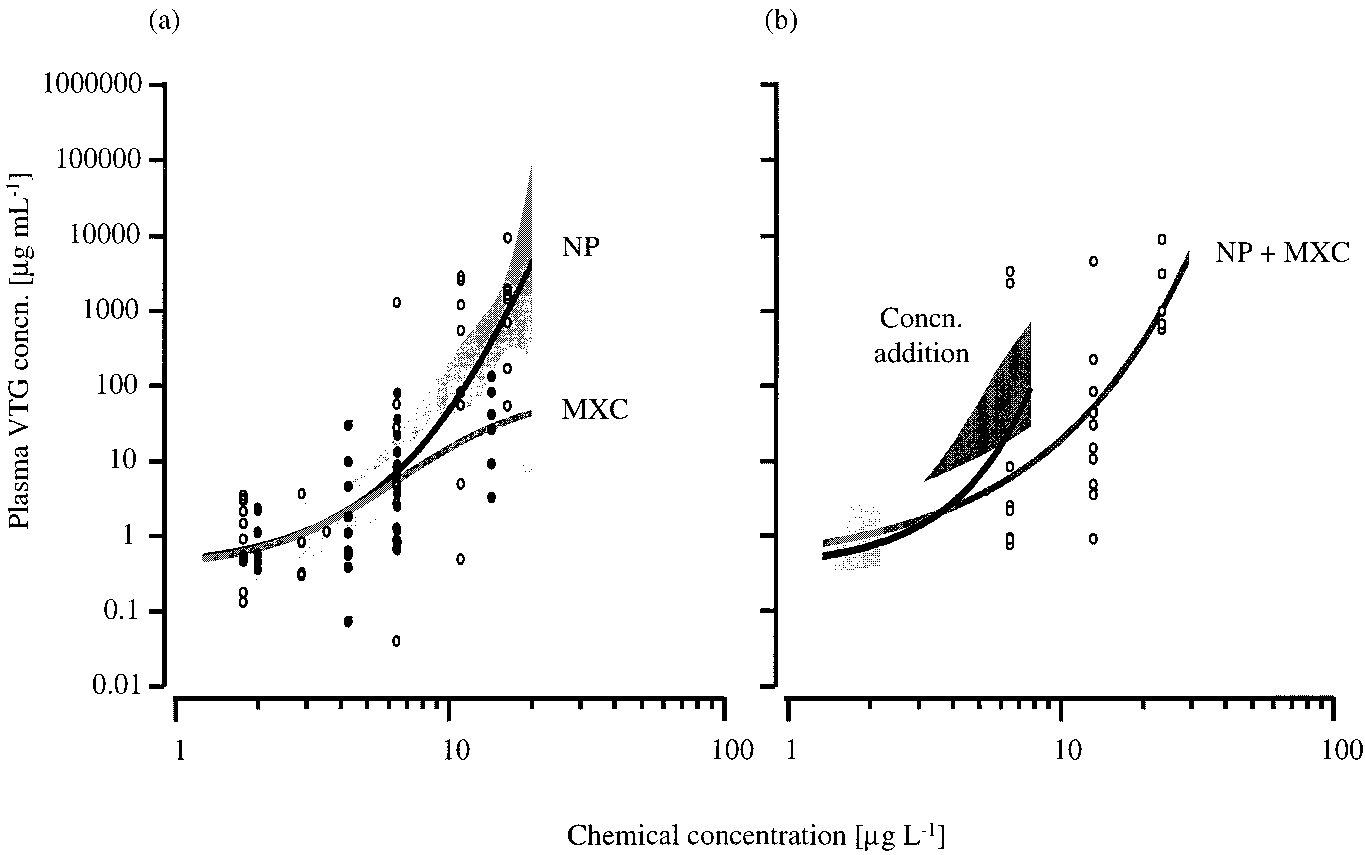 FIGURE 2. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles)
FIGURE 2. Plasma vitellogenin (VTG) concentrations in female juvenile rainbow trout exposed to (a) estradiol-17 (E2) (closed circles)