Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Signature i-86.indd
Breeding cowpea for resistance to insect pests: attempted crosses between cowpea and Vigna vexillata Abstract Cowpea is grown mainly for its protein-rich grains, which is consumed in various forms in sub-Saharan Africa. Average grain yield in farmers’ fi elds is generally low due to a number of biotic and abiotic stresses. The most important of the biotic stress factors causing extensive grain yield losses in cowpea are postfl owering insect pests such as the legume pod borer and pod sucking bugs. Availability of varieties with resistance to these pests will be attractive to cowpea farmers as the crop could then be grown with less dependence on expensive, often adulterated chemicals that are not particularly environmentally friendly. To be able to develop such varieties, it is necessary that genes conferring resistance to these pests are available in the cowpea genome. Genes conferring resistance to these pests were found to exist in the genomes of some wild Vigna species such as V. vexillata and V. oblongifolia and efforts were made to transfer these genes from the wild Vigna sp. to cowpea. Pods were retained for up to seven days after pollination when V. vexillata lines served as female parents with cowpea. Embryos in pods resulting from these crosses did not develop beyond the globular stage. Several procedures aimed at overcoming this incompatibility were adopted without success. Among the techniques used to overcome incompatibility were in vitro culture of interspe- cifi c hybrid embryos, hormonal treatments of fl ower buds prior to pollination, and polyploidization. No interspecifi c hybrids were obtained following the several attempts made, thus suggesting that very strong cross-incompatibility exists between cowpea and V. vexillata. Introduction Cowpea (Vigna unguiculata [L.] Walp.) is grown mainly for its grain, which contains between 22 and 32% protein on a dry weight basis. The grain is one of the cheapest sources of protein in the diets of peoples of West and Central Africa where cowpea is also an important crop. The dried grain is consumed after being processed into different food forms while the haulms from dried and shelled pods as well as fodder, are a good source of quality feed for livestock. Farmers in the dry savanna areas of West and Central Africa derive some income from selling cowpea fodder to livestock owners, particularly during the dry season.
Every stage in the life cycle of cowpea has at least one major insect pest that could
cause serious damage and impact yield negatively. When postfl owering insect pests infest
1. International Institute of Tropical Agriculture, PMB 5320, Oyo Road, Ibadan, Nigeria. Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
cowpea fi elds and cause heavy damage to grain yield, farmers, especially those in the dry savanna area, resort to harvesting the fodder in order to get some income. There is, however, no doubt that farmers get more fi nancial benefi t from cowpea grain than from fodder. In order for farmers to obtain high grain yield from their cowpea fi elds it is nec-essary for them to spray the cowpea plants with insecticides a number of times. Until recently, cowpea required being sprayed with insecticides up to fi ve times or even more if high grain yield was to be obtained. Relatively high grain yield can now be obtained with two or three insecticide sprays. The high grain yield which can now be obtained with fewer insecticide spray regimes can be attributed to progress that has been made through genetic improvement whereby genes for resistance to some diseases and prefl owering insect pests such as aphids have been incorporated into new cowpea varieties. Also, there are cowpea lines that combine these resistance genes with low levels of resistance to the fl ower bud thrips (Megalurothrips sjostedti). Even some traditional farmer varieties have also been improved by introgressing into them these generally simply inherited resistance genes. Furthermore, the population dynamics for most of the insects have been studied and information obtained has been found useful to target the time of intervention by farmers. No cowpea line has been identifi ed as possessing the desired levels of resistance to the legume pod borer (Maruca vitrata) and pod sucking bugs (Clavigralla tomentosicollis, Anoplocnemis curvipes, and Riptortus dentipes) all of which are postfl owering pests. The legume pod borer and pod sucking bugs can cause tremendous grain-yield losses in cowpea if appropriate control measures are not taken.
The most economical and environmentally friendly way of controlling these insect
pests would be through host-plant resistance. Introgressing genes for resistance to the insect pests into cowpea should result in the availability of varieties which can be grown by farmers in sub-Saharan Africa with minimal use of chemicals. This will lead to a reduction in the cost of cowpea production, thereby increasing the profi t margin for farmers. Essentially cowpea production will become more attractive to the generally resource-poor farmers in the savanna zones of Africa. In addition, the farmers would be healthier as they no longer need to handle toxic chemicals while at the same time pollution of their environment would be immensely reduced. Because of the poten-tially immense benefi ts of growing insect-resistant cowpea varieties, no efforts should be spared in the search for and transfer of the desired genes from landraces and wild Vigna species to cultivated cowpea. To this end, a wide range of accessions from the cowpea germplasm collection as well as those of its wild and weedy cross-compatible relatives was screened in order to identify those with genes for resistance to the pests that wreak havoc on cowpea production. None of the tested cultivated cowpea lines and their cross-compatible wild relatives showed the desired high level of resistance to these pests. Several accessions of some Vigna species, such as those belonging to V. vexillata, V. davyi, V. oblongifolia, and V. luteola, were also screened, among others, for resistance to insect pests of cowpea. The results showed that some accessions of V. vexillata and V. oblongifolia have good levels of resistance to the insect pests that devastate cowpea. The Vigna species whose accessions showed resistance to the major postfl owering insect pests of cowpea do not belong to same primary or secondary gene pool as cowpea and this could constitute a major constraint to moving the desirable genes into cultivated cowpea varieties.
A phylogenetic study that was carried out involving various Vigna species and based
on RFLP markers indicated that among species showing high levels of resistance to the
insect pests, V. vexillata is the closest to cowpea (V. unguiculata) (Fatokun et al. 1993). In this same study, a wild and cross-compatible relative of cowpea V. unguiculata ssp. dekindtiana var. pubescens was linked to V. vexillata when the various accessions of Vigna species tested were displayedon a minimum spanning tree.
The various accessions of V. vexillata showed high levels of resistance to pod sucking
bugs, fl ower thrips, Marucavitrata, bruchid, and Striga gesnerioides among others. Pos-session of these traits makes interspecies crosses between it and cowpea very attractive and worth pursuing. Hence, crosses were initiated between cowpea and V. vexillata withthe aim of transferring the genes conferring resistance to insect pests from the latter to cowpea. While making these crosses (V. vexillata × cowpea) it was observed that some pods were retained for up to seven days or even more when cowpea is the pollen parent. However, in the reciprocal crosses pods were not retained as emasculated fl owers drop within one day following cross pollination. On the other hand, the pods that were retained by pollinating V. vexillata with cowpea developed slowly as the seeds contained therein. By the time these pods attained their maximum size, they only approximated the size attained by four-day-old pods resulting from selfi ng. In all the crosses, fl owers were emasculated and pollinated a day before anthesis. This was to ensure that pollen tubes reached the ovule in order to release the male (sperm) nuclei in time for fertilization to take place. No viable interspecifi c hybrid seed was obtained from any of the several hundreds of crosses made, thus suggesting a strong cross incompatibility between the two species, V. vexillata and V. unguiculata.Overcoming interspecies incompatibility There are a number of procedures that have been used by breeders to overcome barriers that prevent gene exchange between distantly related plant species. These have been used to successfully effect interspecies hybridization in several crops. Among the procedures commonly used are making reciprocal crosses (Thomas and Waines 1982), crossing between different accessions of both species (Harlan and de Wet 1977), polyploidization followed by crossing, polyploidization of the F interspecies hybrid (where the F is sterile),
embryo rescue (Przywara et al. 1989), bridging crosses (Hermsen and Ramanna 1973), and hormonal treatment of fl ower buds prior to or after pollination (Larter and Chaubey 1965; Sitch and Snape 1987), among others. Some of the methods used to overcome constraints to gene exchange through wide crossing in several crops were tried in the attempted cross between cowpea and V.vexillata and these are reported in the following sections. Crossing several accessions of both species: Reports of previous wide crossing activities in some crops have shown that hybrids between certain accessions of a species are more productive than others. This is because certain accessions of a species are better combin- ers with some other individuals of another species. In tobacco (Nicotiana tabacum) Pit- tarelli and Stavely (1975) observed that when three different cultivars were crossed to N. repanda only one combination gave F hybrid plants. Harlan and de Wet (1977) also tested
a number of Tripsacumdactyloides in combination with corn and found that only one of the T. dactyloides accessions was effective in transferring genetic material to maize. In the attempted cross between cowpea and V. vexillata, some pods are retained on V. vexillata when emasculated fl owers are pollinated with cowpea but none were retained in reciprocal crosses. It is conceivable that not all lines of V. vexillata will respond in the same way as, for example, in the frequency of pod retention when fl owers are pollinated with different
Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
cowpea accessions. It is also possible that in some specifi c combinations the embryos may develop beyond the globular stage. Perhaps some V. vexillata lines might even support the development of pods with well-formed seeds to maturity while others do not. Hence several accessions of V. vexillata were selected for crossing with cowpea.
Pollen from four wild cowpea relatives belonging to V. unguiculata ssp. dekindtiana and
ten cultivated cowpea (V. unguiculata) lines were used to pollinate emasculated fl owers of 64 different accessions of V. vexillata. There were differences among the accessions of V. vexillata used in making these crosses in the frequency of pods they retained fol-lowing pollination with cowpea lines or wild cowpea relatives. While accession TVNu 73 retained up to 70% pods following the interspecies pollination, only a few pods were retained by some other accessions such as TVNu 1359 (Table 1). It should be noted that the retained pods were on the plants for no longer than eight to ten days after pollination. They shriveled and fell off the plant prematurely. Pods resulting from selfi ng on V. vexil-lata remain on the parent plants until they dry and are harvested. No appreciable differ-ences were observed in the frequency of pod retention on the basis of which cowpea or dekindtiana line was used as pollen parent. Also, there were no observed differences in embryo development when random samples of ovules in retained pods were dissected. Essentially none of the ovules from the interspecifi c hybridization had an embryo that developed beyond the globular stage. Use of mixed pollen: Cowpea pollen grains do produce tubes albeit at low frequencies when placed on the stigma of V. vexillata. Also, some of the developed pollen tubes are malformed and are therefore unable to penetrate the style fast enough to reach the ovule in order to effect fertilization (Barone and Ng 1990). A few pollen grains of the female (V. vexillata) plants were deliberately placed on the stigma along with some of cowpea. Pods developed on the vexillata plants when the mixed pollen grains were used. The number of normal sized seeds in each pod was few but none of the seeds resulted from interspecifi c hybridization. Payan and Martin (1975) used the mixed pollen technique to successfully effect interspecies cross in the genus Passifl ora (passion fruit). Application of growth hormones: Growth promoting hormones have been used to facili- tate interspecies crosses in many crops. Generally, hormones are known to prolong the period during which fruits are retained on plants. In Phaseolus, Al-Yasiri and Coyne (1964) used growth hormones to prolong the period of pod retention following wide crossing to 30 days as against 15 for untreated pods. Gibberellic acid and NAA are two commonly Table 1. Frequency of pod retention by some accessions of Vigna vexillata following pollination with cowpea.
used hormones to treat fl owers in order to enhance interspecies crosses. Two auxins (2,4-D & NAA) and one cytokinin (kinetin) were applied as sprays at low concentrations (approximately 1.0 mg/l) and in various combinations on fl owers of V. vexillata before or after pollination with cowpea. In particular, 2,4-D was effective in promoting the reten-tion of V. vexillata fl owers pollinated with cowpea and subsequently the pods resulting from the cross-pollination. Pods that formed from V.vexillata fl owers sprayed with 2,4-D and pollinated with cowpea developed on the plants and at maturity were bigger in size than those resulting from selfi ng with no 2,4-D sprayed (Fig. 1). These pods remained on the peduncles until they dried as for normal pods resulting from self-pollination. When pods resulting from fl owers sprayed with 2,4-D had matured and were opened, the ovules contained in them did not develop beyond the size of three-day-old ovules of selfed pods. In addition, there was a mass of white colored loose callus-like structures, which fi lled the spaces between adjacent ovules (Fig. 2). When emasculated fl owers were sprayed with 2,4-D but not pollinated, the fl owers remained attached to the peduncle for up to six days before falling off. Pods were, however, not initiated from such nonpollinated fl owers even with the hormonal treatment. The retention and development of pods on V. vexillata following pollination with cowpea and 2,4-D spray is further evidence that fertilization does occur, leading to embryo initiation. According to Barone and Ng (1990), between 15 and 20% of ovules are fertilized when V. vexillata fl owers are pollinated with cowpea. However, the embryos in ovules could not go through the normal stages of development for some reasons. These observations show that prolonging the retention and develop-ment of pods resulting from the V. vexillata by cowpea crosses on the female parent did not lead to further embryo development. Deakin et al. (1971) made similar observations in interspecies crosses in cucumber.
The application of NAA as spray to fl ower buds was not as effective as 2,4-D in promot-
ing retention of pods on V.vexillata following pollination with cowpea. The pods result-ing from fl owers sprayed with NAA increased in size and were only slightly bigger than those that were not sprayed. Also NAA did not increase the frequency of pod retention as compared to when cross-pollinated fl owers were not treated with a hormone. Embryo rescue: Developments and improvements in tissue and cell culture techniques have contributed immensely to progress made in the exchange of genes between species in many crops. In vitro culture methods have been used to rescue young interspecifi c hybrid embryos prior to their abortion. This is particularly important in situations where the cause of incompatibility occurs postfertilization such as endosperm abortion and eventual starvation of the embryo. Fatokun and Singh (1987) needed to rescue embryos of the cross between cowpea and a wild relative, V. unguiculata ssp. pubescens, otherwise the embryos resulting from the cross collapsed before attaining full development. Barone et al. (1992) reported that the embryo and endosperm resulting from the cross between V.vexillata and cowpea collapsed within fi ve and eight days following pollination. The development of an embryo especially during the early stages depends on the existence of a well-formed endosperm, which is the primary source of nourishment for the embryo. Also, it is essential that a harmonious relationship should exist between the embryo and endosperm tissue if the former is to go through the process of development.
When excised, the embryos in ovules resulting from pollinating V.vexillata with
cowpea attained the globular stage of development (Fatokun 1991). The successful rescue of interspecifi c hybrid embryos that are this young (i.e., at the globular stage) has been
Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
Figure 1. Pods of Vigna vexillata; (A) mature selfed pods, (B) mature pods from fl owers pollinated with cowpea and sprayed with 2,4-D, and (C) seven-day-old pods from fl owers pollinated with cowpea but not treated with 2,4-D. Figure 2. Opened pod of Vigna vexillata from fl owers pollinated with cowpea and sprayed with 2,4-D showing ovules (arrows) which did not develop into seeds.
diffi cult to achieve in many plants. Embryo rescue is more successful as the embryo gets older. Usually the development of embryos into plants occurs more readily once they have passed the globular stage of development and beyond. It is obvious from the observations and reports mentioned above that fertilization does take place when pollen of cowpea are placed on the stigma of V. vexillata.
Entire ovules resulting from pollinating V. vexillata with cowpea were cultured in
MS media containing 10% coconut water, 1% casein hydrolysate, and varying levels and combinations of sucrose and benzyl adenine. In ovulo culture was used because the embryos are small and diffi cult to dissect and excise. The presence of low levels (1–3%) of sucrose along with the other organic components added to the media encouraged the development of young selfed embryos (as young as four days) of both species to develop into plants in the culture tube. However, none of the hybrid embryos developed into plants following placement in the culture media. A few of the ovules (both selfed and hybrid) formed calluses especially in the media containing growth hormone, but when subcultured, no plants could be regenerated from them.
Polyploidization:Increasing chromosome number of one or both species can enhance crossability between two species. This is particularly so in cases where the two species being crossed differ in their genome number. However, both cowpea and V. vexillata have the same number of chromosomes (2N = 22) as all other members of the genus Vigna with the exception of V. glabrescens which has 2N = 2X = 44. Vigna glabrescens is the only naturally occurring polyploid in the subtribe Phaseolinae (Marechal et al. 1978). Polyploids were induced in cowpea following treatment of shoot tips of young seedlings with a weak solution of colchicine. Different accessions responded differently to colchicine such that a higher frequency of polyploids were induced in some than in others following similar treatments. The induced polyploids are fertile but produce fewer seeds per pod compared to their diploid counterparts. In addition the plants produced only a few pods each. The polyploid cowpea plants were characterized by thick leaves with large guard cells around the stomates, larger fl owers, and pollen grains that were mostly rectangular in shape. Root tip cells obtained from the plants had 2N = 2X = 44 chromosomes. Young seedlings of V. vexillata, treated with colchicine were more sensitive to the chemical than cowpea. Further growth and development of seedlings were arrested at the shoot tips fol- lowing application of 0.1% colchicine for a period of 12 hours. Application of colchicine at 0.1% to seedling shoot tips for 24 hours was found to be most effective in cowpea. In V. vexillata seedlings treated with colchicine new shoots developed from the roots rather than from the shoot tips. In an attempt to promote development of new shoots from treated buds of V. vexillata, young shoots were grafted on cowpea plants as stock. Shoot tips and axillary buds of the scion were treated with 0.1% colchicine for 12 hours. No shoots developed from any of the axillary buds or shoot tips of the scion treated with colchicine. Hence no polyploids could be induced in V. vexillata using the same concentration of colchicine that was effective on cowpea. Pollen of V. vexillata was placed on the stigma of polyploid cowpea fl owers but all such fl owers dropped within 24 hours of pollination hence no pods developed on the polyploid cowpea plants. Bridge crossing: Successful interspecifi c crosses have been made in plants through bridg- ing of crosses. Where direct crosses are not feasible between two species their genomes can be brought together by indirect means. For example Nicotiana tabacum does not readily cross with N. repanda. However, both species can cross successfully with N. sylvestris. Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
The desirable gene for disease resistance present in N. repanda could be transferred to tobacco by fi rst crossing N. repanda to N. sylvestris and the progeny of this cross was then crossed to tobacco (Burk 1967). In order to effect gene transfer from V. vexillata to cowpea, crosses were made between the former and a close relative V. davyi on the one hand and between cowpea and V. unguiculata ssp. dekindtiana on the other. Both V. vexillata and V. davyi belong to the same section Plectotropis in the genus Vigna and this was the fi rst reported successful cross between V. vexillata and any other Vigna species. However, V. davyi and V. unguiculata ssp. dekindtiana could not be crossed successfully. Cowpea and V. unguiculata ssp. dekindtiana belong to the same section Catiang of the genus. The hybrid resulting from the cross between V.vexillata and V. davyi was partially fertile (Table 2) as that between cowpea and its wild relative. The hybrids (V. vexillata × V. davyi and V. unguiculata × V. unguiculata ssp. dekindtiana) were crossed to each other and to the four parents but the efforts did not yield the desired products as no seeds were set in the crosses between members from different sections.
Use of a parthenocarpic cowpea line: A cowpea line (RI 36) showing parthenocarpy was identifi ed among the progeny of a cross between IT84s-2049 and IT88s-524-B at the University of California, Riverside (J. Ehlers, personal communication). This cowpea line has the capacity to form and retain pods to maturity from emasculated fl owers even when not pollinated. This cowpea line therefore served as the female parent and pollinated using a number of V. vexillata accessions. Seeds formed in the pods and these appeared to develop normally for the fi rst ten days after which they started to shrivel. There was a mass of cells connecting each seed to the pod wall. Seeds were excised from pods on different days after pollination for placement in the culture media. Embryos could not be readily distinguished in the seed hence all seeds (ovules) were excised and placed in culture media. The only development observed on the cultured seeds was root initiation (Fig. 3) but no shoots were formed. Table 2. Morphological attributes of F interspecifi c hybrid between Vigna vexillata and V. davyi and their parents.
†V. davyi. ‡V. vexillata. Conclusion Attempts have been made to cross cowpea with V. vexillata using various techniques that have successfully been used to effect wide crosses in some other crops. These efforts did not yield the expected results, thus suggesting the existence of a strong cross incompatibility barrier between cowpea and V. vexillata. The causes of incompatibility between the two species are both pre- and postfertilization. In the fi rst place, only a few Figure 3. Root developing from cultured seed of cowpea line RI 36 following pollination with Vigna vexillata.
cowpea pollen tubes are able to penetrate the styles of V. vexillata and reach the ovule in order to effect fertilization. There is suffi cient evidence that fertilization does occur, albeit at relatively low frequency. The fertilization probably gives rise to diploid zygotes, which develop to the globular stage embryo. The nondevelopment of the hybrid embryos beyond the globular stage may be an indication of incomplete fertlization in which the second male nucleus does not fuse with the diploid endosperm nucleus to give the triploid tissue that normally feeds the embryo. Consequently there is no triploid endosperm tissue formed following the cross between cowpea and V. vexillata. The absence of the endosperm leads to starvation and subsequent collapse of the embryos.
There is in the genome of V. vexillata a repertoire of genes that could confer resistance
to several of the pests and diseases to which cowpea succumbs. The attempts made so far, using sexual means, to move desirable genes from V. vexillata to cowpea have not yielded the desired results. Perhaps other avenues by which these genes could be accessed should be explored. An approach would be the identifi cation and cloning of these genes which eventually could be used to transform cowpea. This is a much longer route to take but it might be worth the efforts because of the potential benefi t.
References
Al-Yasiri, A. and D.P. Coyne. 1964. Effects of growth regulators in delaying pod abscission and
embryo abortion in the interspecifi c cross Phaseolus vulgaris × P. acutifolius. Crop Science 4: 433–435.
Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
Barone, A., A. Del Guidice, and N.Q. Ng. 1992. Barriers to interspecifi c hybridisation between
Vigna unguiculata and V. vexillata. Sexual Plant Reproduction 5: 195–200.
Barone, A. and N.Q. Ng. 1990. Embryological study of crosses between Vigna unguiculata and
V. vexillata. Pages 151–160 in Cowpea genetic resources, edited by N.Q. Ng and L.M. Monti. International Institute of Tropical Agriculture, Ibadan, Nigeria.
Burk, L.G. 1967. An iterspecifi c bridge-cross—Nicotiana repanda through N. sylvestris to N. taba-cum. Journal of Heredity 58: 215–218.
Deakin, J.R., G.W. Bohn, and T.W. Whitaker. 1971. Interspecifi c hybridisation in Cucumis.
Econonomics of Botany 25: 195–210.
Fatokun, C.A. 1991. Wide hybridisation in cowpea: problems and prospects. Euphytica 54: 137–
Fatokun, C.A., D. Danesh, N.D. Young, and E.L. Stewart. 1993. Molecular taxonomic relationships
in the genus Vigna based on RFLP analysis. Theoretical and Applied Genetics 86: 97–104.
Fatokun, C.A. and B.B. Singh. 1987. Interspecifi c hybridisation between Vignapubescens and
V. unguiculata (L.) Walp. through embryo rescue. Plant Cell, Tissue, and Organ Culture 9: 229–233.
Harlan, J.R. and J.M.J. de Wet. 1977. Pathways of genetic transfer from Tripsacum to Zea mays.
Proceedings of the National Academy of Science, USA 74: 3494–3497.
Hermsen, J.G. and M.S. Ramanna. 1973. Double bridge hybrids of Solanum bulbocastanum and
cultivar S. tuberosum. Euphytica 22: 457–466.
Larter, E. and C. Chaubey. 1965. Use of exogenous growth substances in promoting pollen tube
growth and fertilisation in barley-rye cross. Canadian Journal of Genetic Cytology 7: 511–518.
Marechal, R., J.M. Mascherpa, and F. Stainer. 1978. Etude taxonomique d’un groupe complexe
d’espèces des genres Phaseolus et Vigna (Papillionaceae) sur la base de données morphologiques, et polliniques traitées par l’analyse informatique. Boissiera 28: 1–273.
Payan, F.R. and F.W. Martin. 1975. Barriers to the hybridisation of Passifl ora species. Euphytica
Pittarelli, G.W. and J.R. Stavely. 1975. Direct hybridisation of Nicotianarependa × N. tabacum.
Przywara, L., D.W.R. White, P.M. Sanders, and D. Maher. 1989. Interspecifi c hybridisation of
Trifolium repens with T. hybridum using in ovulo embryo culture. Annals of Botany 64: 613–624.
Sitch, L.A. and J.W. Snape. 1987. Factors affecting haploid production in wheat using the Hordeum bulbosum system. 1. Genotype and environmental effects on pollen grain germination, pollen tube growth and the frequency of fertilisation. Euphytica 36: 483–496.
Thomas, C.V. and J.G. Waines. 1982. Interspecifi c hybrids between Phaseolus vulgaris L. and P. acutifolius: Field trials. Annual Report of Bean Improvement Cooperative 25: 58–59.
inzwischen festgestellt haben werden, laufen alle relevanten Themen nach ähnlichen Mustern ab, wobei man zwangsläufig nicht umhin kommt, an gewisse Verschwörungstheorien zu denken. Ich bin heute zu der Meinung gekommen, dass es gar keine Ver-schwörungstheorien gibt. Es handelt sich bei näherem Hinschauen um recht handfeste Verschwörungen, um Tatsachen, völlig ohne Theorien, mit denen wir of
CAGE CULTURE OF SOUTH AMERICAN CATFISH (RHAMDIA SAPO) PRELIMINARY RESULTS IN THE SALTO GRANDE RESERVOIR (ARGENTINA) LAURA M. LUCHINI and ROLANDO QUIROS Instituto Nacional de Investigación y Desarrollo Pesquero (INIDEP) C.C. 175, 7600 Mar Del Plata. Argentina The first experimental cage culture of South American catfish (Rhamdia sapo) was carried out in the Salto Grande

 Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
Figure 1. Pods of Vigna vexillata; (A) mature selfed pods, (B) mature pods from fl owers
Breeding for resistance to insect pests: crosses between cowpea and Vigna vexillata
Figure 1. Pods of Vigna vexillata; (A) mature selfed pods, (B) mature pods from fl owers 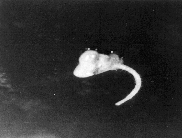 Figure 3. Root developing from cultured seed of cowpea line RI 36 following pollination
Figure 3. Root developing from cultured seed of cowpea line RI 36 following pollination