Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
No job name
Chem. Res. Toxicol. 2006, 19, 164-172 The Greater Reactivity of Estradiol-3,4-quinone vs Estradiol-2,3-quinone with DNA in the Formation of Depurinating Adducts: Implications for Tumor-Initiating Activity
Muhammad Zahid, Ekta Kohli, Muhammad Saeed, Eleanor Rogan, and Ercole Cavalieri*
Eppley Institute for Research in Cancer and Allied Diseases, UniVersity of Nebraska Medical Center,986805 Nebraska Medical Center, Omaha, Nebraska 68198-6805
Strong evidence supports the idea that specific metabolites of estrogens, mainly catechol estrogen-
3,4-quinones, can react with DNA to become endogenous initiators of breast, prostate, and other humancancers. Oxidation of the catechol estrogen metabolites 4-hydroxyestradiol (4-OHE2) and 2-OHE2 leadsto the quinones, estradiol-3,4-quinone (E2-3,4-Q) and estradiol-2,3-quinone (E2-2,3-Q), respectively. Thereaction of E2-3,4-Q with DNA affords predominantly the depurinating adducts 4-OHE2-1-N3Ade and4-OHE2-1-N7Gua, whereas the reaction of E2-2,3-Q with DNA yields the newly synthesized depurinatingadduct 2-OHE2-6-N3Ade. The N3Ade adducts are lost from DNA by rapid depurination, while the N7Guaadduct is lost from DNA with a half-life of ∼3 h at 37 °C. To compare the relative reactivity of E2-3,4-Qand E2-2,3-Q, the compounds were reacted individually with DNA for 0.5-20 h at 37 °C, as well as inmixtures (3:1, 1:1, 1:3, and 5:95) for 10 h at 37 °C. Depurinating and stable adducts were analyzed. Insimilar experiments, the relative reactivity of 4-OHE2 and 2-OHE2 with DNA was determined afteractivation by lactoperoxidase, tyrosinase, prostaglandin H synthase (PHS), or 3-methylcholanthrene-induced rat liver microsomes. Starting with the quinones, the levels of depurinating adducts formed fromE2-3,4-Q were much higher than that of the depurinating adduct from E2-2,3-Q. Similar results wereobtained with lactoperoxidase or tyrosinase-catalyzed oxidation of 4-OHE2 and 2-OHE2, whereas withactivation by PHS or microsomes, a relatively higher amount of the depurinating adduct from E2-2,3-Qwas detected. These results demonstrate that the E2-3,4-Q is much more reactive with DNA than E2-2,3-Q. The relative reactivities of E2-3,4-Q and E2-2,3-Q to form depurinating adducts correlate with thecarcinogenicity, mutagenicity, and cell-transforming activity of their precursors, the catechol estrogens4-OHE2 and 2-OHE2. This is essential information for understanding the cancer risk posed by oxidationof the two catechol estrogens. Introduction
Catechol estrogens, 2-hydroxyestrone(estradiol) [2-OHE1(E2)]
The initial failure to demonstrate that estrogens induce
1(E2), are among the major metabolites of E1 and
mutations in bacterial and mammalian test systems (1-6)
2. If these metabolites are oxidized to the electrophilic catechol
estrogen quinones, they may react with DNA. The 4-catechol
resulted in the classification of estrone (E1)1 and estradiol (E2)
estrogens are carcinogenic in Syrian golden hamsters, as well
as epigenetic carcinogens that function mainly by stimulating
as CD-1 mice (1, 20, 21), whereas the 2-catechol estrogens are
abnormal cell proliferation via estrogen receptor-mediated
not carcinogenic in the hamsters (1, 20) and are borderline
processes (3, 7-12). The stimulated cell proliferation can result
carcinogens in CD-1 mice (21).
in more accumulation of genetic damage leading to carcino-
genesis (9, 10, 12).
1(E2) is easily oxidized to catechol estrogen-3,4-
Compelling strong evidence has resulted in a new paradigm
predominantly depurinating adducts (13-16). These adducts
of cancer initiation by estrogens. Discovery that specific
generate apurinic sites that may lead to cancer-initiating
oxidative metabolites of estrogens can react with DNA (13-
mutations (17-19), which transform cells (22-25), thereby
16) led to and supported the hypothesis that estrogen metabolites
can become endogenous chemical carcinogens by generatingmutations (17-19) that can lead to initiation of cancer. The
To determine the DNA adducts of E1(E2)-3,4-Q, standard
initiating mechanism can occur in hormone-dependent and
adducts were synthesized by reaction of the quinones with
deoxyguanosine (dG), deoxyadenosine (dA), and the nucleobaseAde (16, 26). The reaction of E1(E2)-3,4-Q with dG affordedthe depurinating adduct 4-OHE1(E2)-1-N7guanine (Gua) by 1,4-
* To whom correspondence should be addressed. Tel: 402-559-7237.
Fax: 402-559-8068. E-mail: ecavalie@unmc.edu.
Michael addition (26). The reaction of E1(E2)-3,4-Q with dA
1 Abbreviations: Ade, adenine; COSY, chemical shift correlation
did not produce any adduct. However, the reaction of these
spectroscopy; dA, deoxyadenosine; dG, deoxyguanosine; E1, estrone; E2,
quinones with Ade resulted in the formation of 4-OHE1(E2)-1-
estradiol; E2-2,3-Q, estradiol-2,3-quinone; E2-3,4-Q, estradiol-3,4-quinone;
N3Ade by 1,4-Michael addition (16). The rationale for the
FAB-MS, fast atom bombardment tandem mass spectrometry; Gua, guanine;HMBC, heteronuclear multiple bond correlation; HSQC, heteronuclear single
formation of N3Ade adducts is described in the Results and
quantum coherence; IBX, 2′-iodoxybenzoic acid; LP, lactoperoxidase; MC,
3-methylcholanthrene; NOE, nuclear Overhauser effect; OHE2, hydroxy-
When E1(E2)-2,3-Q was reacted with dG or dA, a different
estradiol; PDA, photodiode array; PHS, prostaglandin H synthase; TFA,trifluoroacetic acid; TOCSY, total correlation spectroscopy.
profile of adducts from those formed by E1(E2)-3,4-Q was
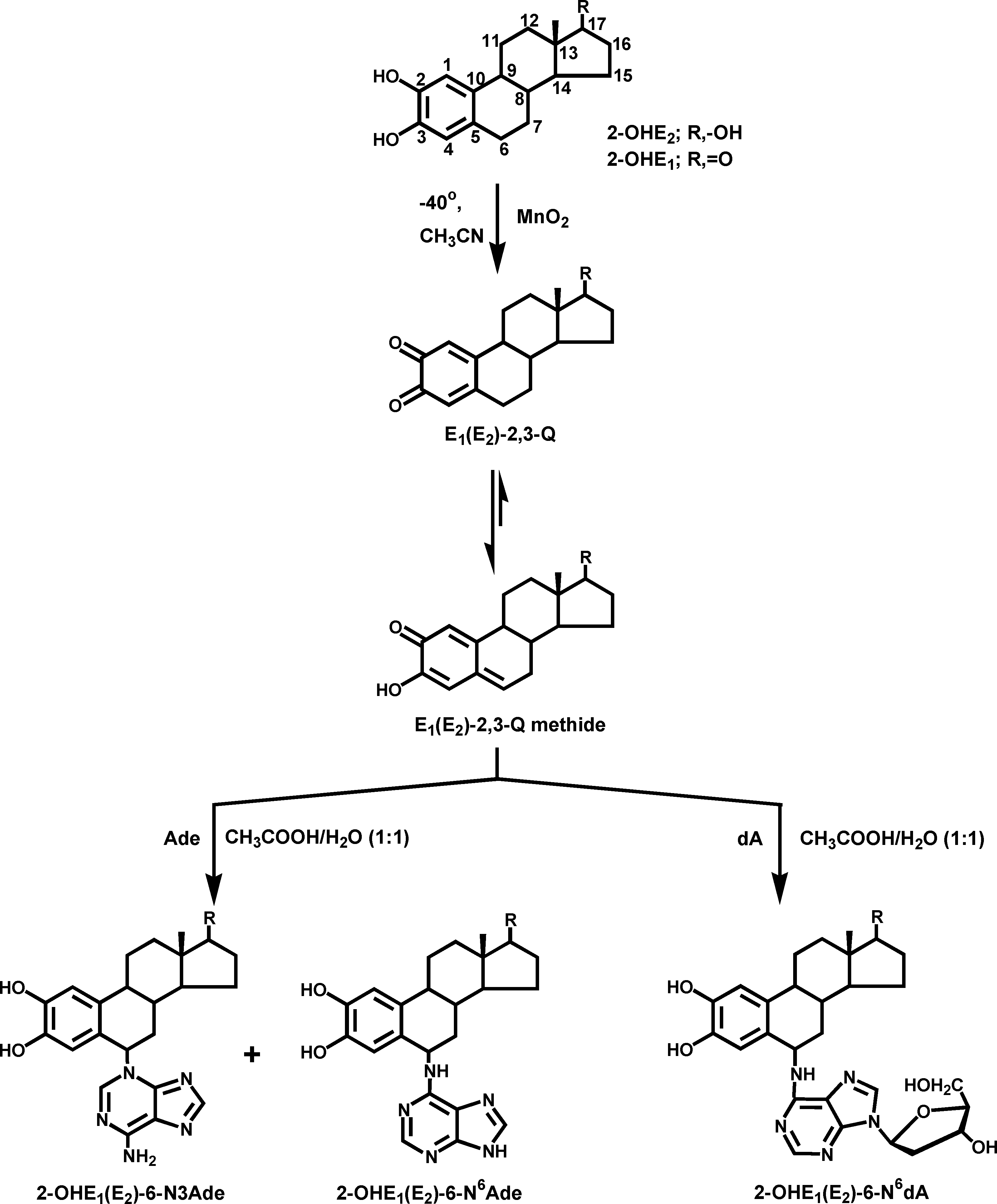
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNAChem. Res. Toxicol., Vol. 19, No. 1, 2006 165 Figure 1. Reaction of E1(E2)-2,3-Q methide with Ade and dA to form the N3Ade and N6Ade adducts and N6dA adducts, respectively.
obtained (26). Reaction of E1(E2)-2,3-Q with dA afforded
Materials and Methods
2-OHE1(E2)-6-N6Ade (Figure 1) and with dG yielded 2-OHE1-
Caution: 2-OHE2, 4-OHE2, E2-2,3-Q, and E2-3,4-Q are hazard-
(E2)-6-N2dG (26). In these reactions, E1(E2)-2,3-Q did not react
ous chemicals and were handled according to NIH guidelines (27).
as quinones, but as their tautomers, the E1(E2)-2,3-Q methide
Chemicals, Reagents, and Enzymes. 2′-Iodoxybenzoic acid
(Figure 1). The electrophilic C-6 of the quinone methide reacted
(IBX) was synthesized from 2′-iodobenzoic acid as described (28).
with the exocyclic amino group of dA or dG via 1,6-Michael
2-OHE2 and 4-OHE2 were synthesized by reacting E2 with IBX
addition to yield the N6dA (Figure 1) and N2dG adducts, which
and then separating the mixture of 2-OHE2 and 4-OHE2 by HPLC.
retain the deoxyribose moiety and are referred to as stable
4-OHE2-1-N3Ade and 4-OHE2-1-N7Gua were synthesized bypublished procedures (16, 26). MnO
adducts because they remain in DNA unless repaired.
ascorbic acid, ammonium acetate, formic acid, sodium phosphate,
In this article, we report the synthesis of the N3Ade
adenine (Ade), DMSO-d6, CH3CN (HPLC grade), H2O2, lactoper-
depurinating adduct obtained by reaction of E1(E2)-2,3-Q with
oxidase (LP, from bovine milk), NADPH, methemoglobin, and
Ade. We also report a study of the competitive reaction between
tyrosinase (from mushrooms) were purchased from Sigma-Aldrich
Chemical Co. (St. Louis, MO). dG and calf thymus DNA were
purchased from USB (Cleveland, OH). Prostaglandin H synthase
2,3-Q) with DNA, as well as the metabolic oxidation of mixtures
(PHS) and arachidonic acid were purchased from Cayman Chemical
of 4-OHE2 (4-hydroxyestradiol) and 2-OHE2 to their quinones
(Ann Arbor, MI), and CH3OH was purchased from Merck KGaA
by selected enzymes in the presence of DNA to form DNA
(Darmstadt, Germany). 3-Methylcholanthrene (MC)-induced rat
adducts. These results will be discussed in relation to the
liver microsomes were prepared as described earlier (29), containing
mechanism of tumor initiation by estrogens.
40 mg protein/mL, with 11.3 nmol cytochrome P450/mg protein. 166 Chem. Res. Toxicol., Vol. 19, No. 1, 2006
Bond Elute Certify II SPE cartridges were purchased from Varian
the data were acquired and processed using the CoulArray software
package. Peaks were identified by both retention time and peak
Activated MnO2 was prepared as previously described (30), by
height ratios between the dominant peak and the peaks in the two
treating concentrated aqueous KMnO4 with aqueous MnSO4 solu-
adjacent channels. The 2-OHE2-6-N3Ade adduct eluted at 14.47
tion kept at 90 °C, until a slight excess of KMnO4 was present, as
min, 4-OHE2-1-N3Ade eluted at 16.55 min, and 4-OHE2-1-N7Gua
indicated by the pink coloration of the suspension.
eluted at 17.46 min. The depurinating adducts were quantified by
Instrumentation. 1. UV. The UV spectra were obtained during
comparison of peak response ratios with known amounts of
HPLC by using a photodiode array detector (PDA, Waters 996,
Milford, MA) for all synthesized compounds. HPLC separations
Synthesis of Standard Adducts. To a stirred solution of 2-OHE2
or 2-OHE1 (0.2 mmol) in 5 mL of CH3CN at -40 °C, activated
2. NMR. NMR spectra were recorded on a Varian Unity-Inova
MnO2 (2 mmol) was added under an argon atmosphere (Figure 1).
500 instrument operating at a resonance frequency of 499.8 MHz
After 10 min, the yellowish green solution was filtered through a
for 1H and 125.6 MHz for 13C spectra at 25 °C. Samples were
0.45 Gelman acrodisc directly into a flask containing a solution of
dissolved in 600 µL of DMSO-d
signals at 2.5 ppm for 1H and 39.7 ppm for 13C. All two-dimensional
mixture was stirred overnight at room temperature. It was evapo-
(2D) experiments were performed by using the standard Varian
rated to dryness, and the residue was redissolved in dimethyl-
software (VNMR v6.1c). For 2D experiments, relaxation delays
formamide/CH3OH (1:1, 3 mL) and then subjected to preparative
HPLC for purification of the adducts.
2 were recorded for a spectral width of 8000 Hz in
2-6-N3Ade. Yield 43%. UV:
two dimensions. 1H-1H correlations were recorded using correlation
NMR (ppm): 8.93 (s, 1H, OH, exchangeable with D2O), 8.77 (s,
spectroscopy (COSY) and total correlation spectroscopy (TOCSY)
1H, OH, exchangeable with D2O), 8.17 (s, 1H, 8-H-Ade), 7.47 (s,
experiments. The TOCSY experiment was performed in the states
1H, 2-H-Ade), 7.20 (s, 2H, NH2-Ade, exchangeable with D2O),
TPPI mode with a MLEV17 spin lock at 10 kHz field strength. In
6.80 (s, 1H, 1-H), 6.30 (s, 1H, 4-H), 5.65 (d, 1H, J ) 4.0 Hz,
pulsed field gradient 1H-13C heteronuclear single quantum coher-
6-H), 4.45 (s, 1H, 17 -OH, exchangeable with D2O), 3.49 (t, 1H,
ence (HSQC) and heteronuclear multiple bond correlation (HMBC)
J ) 8.5 Hz, 17R-H), 2.20 (dd, 1H, J )
sequences, delays were optimized for coupling constants around
2.13 (m, 1H), 1.96 (d, 1H, J ) 14.0 Hz), 1.75-1.91 (m, 4H), 1.47-
140 and 8 Hz, respectively. One-dimensional nuclear Overhauser
1.52 (m, 2H), 1.38-1.40 (m, 1H), 1.13-1.28 (m, 3H), 0.61 (s,
(NOE) experiments were recorded in difference mode by subtracting
3H, CH3). 13C NMR (ppm): 156.0, 152.2, 149.3, 145.8, 144.0,
one spectrum with irradiation on resonance from another one
140.1, 132.7, 122.8, 118.8, 116.0, 112.6, 80.0, 51.3, 48.8, 43.3,
without irradiation with a relaxation delay of 5 s.
43.1, 36.6, 33.9, 33.5, 29.9, 25.8, 22.7, 11.3. FAB-MS [M + H]+:
3. Mass Spectrometry. Fast atom bombardment tandem mass
422.2170 calcd for C23H28N5O3; observed, 422.2192.
spectrometry (FAB-MS) was conducted at the Nebraska Center for
2. 2-OHE2-6-N6Ade. Yield 9%. UV: λmax 214, 277 nm. 1H NMR
Mass Spectrometry (University of Nebraska-Lincoln) using a
(ppm): 9.25 (br.s, 2H, OH, exchangeable with D2O), 8.23 (s, 1H,
MicroMass (Manchester, England) AutoSpec high-resolution mag-
8-H-Ade), 8.07 (s, 1H, 2-H-Ade), 7.73 (br.s, 1H, 6-NH-Ade,
netic sector mass spectrometer. The instrument was equipped with
exchangeable with D2O), 6.67 (s, 1H, 1-H), 6.55 (s, 1H, 4-H), 5.42
an orthogonal acceleration time-of-flight serving as the second mass
(br.s, 1H, 6-H), 4.40 (s, 1H, 17 -OH, exchangeable with D2O),
spectrometer. Xenon was admitted to the collision cell at a level to
3.53 (t, 1H, J ) 8.5 Hz, 17R-H), 2.09-2.19 (m, 1H), 1.95-2.04
attenuate the precursor ion signal by 75%. Data acquisition and
(m, 1H), 1.73-1.93 (m, 4H), 1.29-1.58 (m, 4H), 1.06-1.25 (m,
processing were accomplished using OPUS software that was
3H), 0.74 (s, 3H, CH3). FAB-MS [M + H]+: 422.2170 calcd for
provided by the manufacturer (Microcasm). Samples were dissolved
in 5-10 µL of CH3OH; 1 µL aliquots were placed on the sample
3. 2-OHE1-6-N3Ade. Yield 41%. UV: λmax 214, 264 nm. 1H
probe tip along with 1 µL of a 1:1 mixture of glycerol/thioglycerol.
NMR (ppm): 8.97 (s, 1H, OH, exchangeable with D2O), 8.77 (s,
4. HPLC. Preparative HPLC was conducted on a Luna-2 C-18
1H, OH, exchangeable with D2O), 8.17 (s, 1H, 8-H-Ade), 7.58 (s,
column (10 µm, 120 Å, 21.2 mm × 250 mm, Phenomenex,
1H, 2-H-Ade), 7.19 (s, 2H, NH2-Ade, exchangeable with D2O),
Torrance, CA) on a Waters 600E solvent delivery system equipped
6.78 (s, 1H, 1-H), 6.27 (s, 1H, 4-H), 5.70 (d, 1H, J ) 4.5 Hz,
with a 996 PDA by using a linear gradient of 10% CH
6-H), 4.45 (s, 1H, 17 -OH), 2.20 (dd, 1H, J ) 10.5, J ) 1.5 Hz),
[0.4% trifluoroacetic acid (TFA)] for 5 min, followed by a linear
2.11-2.13 (m, 1H), 1.96 (d, 1H, J ) 14.0 Hz), 1.75-1.91 (m,
4H), 1.47-1.52 (m, 2H), 1.38-1.40 (m, 1H), 1.13-1.28 (m, 3H),
3CN over 45 min at a flow rate of 6 mL/min.
Analytical HPLC was conducted on a Waters 2690 (Alliance)
0.61 (s, 3H, CH3). FAB-MS [M + H]+: 420.2025 calcd for
Separations Module equipped with a Waters 996 PDA interfaced
to a Digital Venturis Fx 5100 computer by using a Luna-2 C-18
4. 2-OHE1-6-N6Ade. Yield 7%. UV: λmax 210, 277 nm. 1H NMR
column (5 µm, 120 Å, 250 mm × 4.6 mm, Phenomenex). A linear
(ppm): 9.20 (br.s, 2H, OH, exchangeable with D2O), 8.22 (s, 1H,
gradient of 30% CH3CN/70% H2O (0.4% TFA) to 100% CH3CN
8-H-Ade), 8.04 (s, 1H, 2-H-Ade), 7.79 (br.s, 1H, 6-NH-Ade,
in 45 min at a flow rate of 1 mL/min was used to separate the
exchangeable with D2O), 6.65 (s, 1H, 1-H), 6.54 (s, 1H, 4-H), 5.50
reaction mixtures. The concentrations and purity of quinones were
(br.s, 1H, 6-H), 2.34-2.39 (m, 1H), 2.11-2.13 (m, 1H), 1.94-
checked by using a linear gradient starting from 30% CH3CN/H2O
2.06 (m, 4H), 1.81-1.87 (m, 1H), 1.74-1.77 (m, 1H), 1.58-1.62
(0.4% acetic acid) to 100% CH3CN at a flow rate of 1 mL/min in
(m, 1H), 1.33-1.53 (m, 4H), 0.87 (s, 3H, CH3). FAB-MS [M +
15 min with UV detection at 280 and 432 nm, respectively. The
H]+: 420.2025 calcd for C23H26N5O3; observed, 420.2038.
retention time for E2-3,4-Q was 5.75 min and for E2-2,3-Q, 5.32
Preparation of Quinones. 2-OHE2 (2.5 mg, 8.7 µmol) was
min. Analyses of depurinating adducts were conducted on an HPLC
dissolved in 500 µL of CH3CN and stirred at -40 °C. Then,
system equipped with dual ESA model 580 solvent delivery
activated MnO2 (7.5 mg, 86.2 µmol) was slowly added. After 15
modules, an ESA model 540 autosampler, and a 12 channel
min, the yellowish green solution was filtered through a Gelman
CoulArray electrochemical detector (ESA, Chelmsford, MA). The
acrodisc. Because of the instability of E2-2,3-Q, the quinone in CH3-
two mobile phases used were as follows: A, CH3CN:CH3OH:0.1
CN was directly used in an experiment as quickly as possible.
M HCOONH4 (pH 3.7), 15:5:80; and B, CH3CN:CH3OH:0.1 M
4-OHE2 (2.5 mg, 8.7 µmol) was dissolved in 500 µL of CH3CN
HCOONH4 (pH 3.7), 50:20:30. The linear gradient changed from
and stirred at 0 °C. Then, activated MnO2 (7.5 mg, 86.2 µmol)
100% A to 90% B in 50 min. The serial arrays of 12 coulometric
was slowly added. After 20 min, the solution was filtered through
electrodes were set at potentials of -10, 100, 150, 210, 270, 330,
a Gelman acrodisc and an equal amount of DMSO was added. The
390, 450, 510, 570, 630, and 690 mV. The 50 µL injections were
CH3CN was evaporated in a high vacuum rotavapor using dry ice
carried out on a Phenomenex Luna-2 C-18 column (5 µm, 120 Å,
and acetone in the condenser. After evaporation of CH3CN, the
4.6 mm × 250 mm) at 1 mL/min. The system was controlled, and
quinone in DMSO was used for the experiments. The final
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNAChem. Res. Toxicol., Vol. 19, No. 1, 2006 167 Figure 2. Depurinating adducts formed after reaction of E2-3,4-Q and E2-2,3-Q (1:1) with DNA. After 10 h, the level of stable adducts was <1.0 µmol/mol DNA-P and <0.5% of total adducts. Mixtures containing 0.87 mM E2-3,4-Q + E2-2,3-Q and 3 mM DNA in 0.067 M sodium potassium phosphate, pH 7.0, were incubated at 37 °C. At the indicated times, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in the supernatant were analyzed.
concentration of quinone (either individually or together) in the
removed, and the DNA was analyzed for stable DNA adducts (31).
From the remaining incubation mixture, DNA was precipitated with
Covalent Binding of E2-2,3-Q or E2-3,4-Q to DNA. Freshly
2 volumes of ethanol, and the supernatant was used for the analysis
prepared E2-2,3-Q and E2-3,4-Q (0.87 mM total concentration in
of depurinating adducts, as described above. Control reactions were
0.5 mL of DMSO) individually, as well as in mixtures (Figure 2),
carried out under identical conditions with either no enzyme or no
were mixed with DNA (3 mM in 0.067 M sodium potassium
phosphate buffer, pH 7.0) and incubated at 37 °C for different timeperiods (0.5, 1, 2, 3, 5, 10, 15, and 20 h). At the indicated times,
Results and Discussion
DNA was precipitated with two volumes of ethanol, and thesupernatant, containing depurinating adducts, was concentrated to
Synthesis and Characterization of Standard Depurinating
1 mL under low pressure, extracted by using a vac-Elute system
Adducts. The reaction of E2-2,3-Q with Ade yielded the major
with 1 mL of CH3OH/0.1 M NH4OAc (80:20), and finally analyzed
depurinating adduct, 2-OHE2-6-N3Ade (43%), and a minor
for depurinating adducts by HPLC with an electrochemical detector,
product, 2-OHE2-6-N6Ade (9%) (Figure 1). The reaction of E2-
as described above. The levels of depurinating adducts were
determined by comparing peak heights with known adduct stan-
The reaction with dA or Ade does not occur in ring A of the
dards. Precipitated DNA from a 1 mL aliquot from each experimentwas separated for 32P-postlabeling analysis of stable adducts after
estrogen, because of the weak electrophilicity of positions 1
purification of the DNA (31).
and 4. However, after tautomerization of the E1(E2)-2,3-Q to
Covalent Binding of 2-OHE 2 and 4-OHE2 to DNA. Mixtures
1(E2)-2,3-Q methide, the reaction takes place at C-6 via 1,6-
containing different relative concentrations of 2-OHE2 and 4-OHE2
Michael addition. The N3Ade adduct could not be obtained by
(0.87 mM total concentration, Figures 3-7) were incubated with
reaction of E2-2,3-Q with dA because electrophilic attack of
DNA in the presence of different enzymes, including tyrosinase,
the C-6 of E2-2,3-Q methide at the nucleophilic N-3 group of
LP, PHS, and MC-induced rat liver microsomes. The reaction
dA is hindered by the presence of the deoxyribose moiety bound
volume in each experiment was 10 mL. In the tyrosinase experi-
to the adjacent N-9 in dA. The dA also cannot react with E2-
ments, the mixture containing 3 mM calf thymus DNA in 0.067 M
3,4-Q (3), hexestrol-3′,4′-quinone (32), and polycyclic aromatic
sodium-potassium phosphate (pH 7.0), 0.87 mM 2-OHE2 or
hydrocarbons (33-36) to form N3Ade adducts. With DNA,
4-OHE2 (2.5 mg in 500 µL of DMSO), and 1 mg of enzyme (2577units) was incubated at 37 °C for 2 or 10 h. For the LP-catalyzed
instead, N3Ade adducts are formed with estrogens (16, 32) and
reaction, the mixture containing 3 mM calf thymus DNA, 0.87 mM
aromatic hydrocarbons (37, 38) and rapidly lost by depurination,
because the configuration of the deoxyribose moiety in DNA
2 or 4-OHE2 (2.5 mg/ in 500 µL of DMSO), H2O2 (0.5 mM),
and 1 mg of enzyme (97 units) was incubated at 37 °C for 2 or 10
h. For PHS-catalyzed reactions, the mixture containing 3 mM DNA,
The 1H NMR spectrum of 2-OHE2-6-N3Ade showed two
0.87 mM 2-OHE2 or 4-OHE2 (2.5 mg in 500 µL of DMSO), 1 mL
singlets at 8.93 and 8.77 ppm for the two hydroxy groups in
of methemoglobin (2.95 mg/mL in 75 mM KH2PO4, pH 7.5), 1
the catechol moiety, which were confirmed by D2O exchange.
mL of arachidonic acid (50 mM), and 800 µL of PHS (400 units)
The two single proton integration signals at 8.17 and 7.47 ppm
was incubated at 37 °C for 2 or 10 h. For the microsome-catalyzed
were assigned to H-8 (Ade) and H-2 (Ade), respectively, based
reaction, the 10 mL mixture containing 3 mM DNA in 150 mMTris-HCl (pH 7.5), 150 mM KCl, 5 mM MgCl
on similar values in other N3Ade adducts (16, 32, 34-36). In
fact, the significant upfield shift of the H-2 (Ade) proton at 7.47
2 (2.5 mg in 500 µL of DMSO), 10 mg of microsomal
protein (40 mg/mL), and NADPH (0.6 mM) was incubated at 37
ppm, as compared to H-2 in unsubstituted Ade, strongly
°C for 2 or 10 h. A 1 mL aliquot from each reaction mixture was
indicates that this proton is shielded by the aromatic ring of the
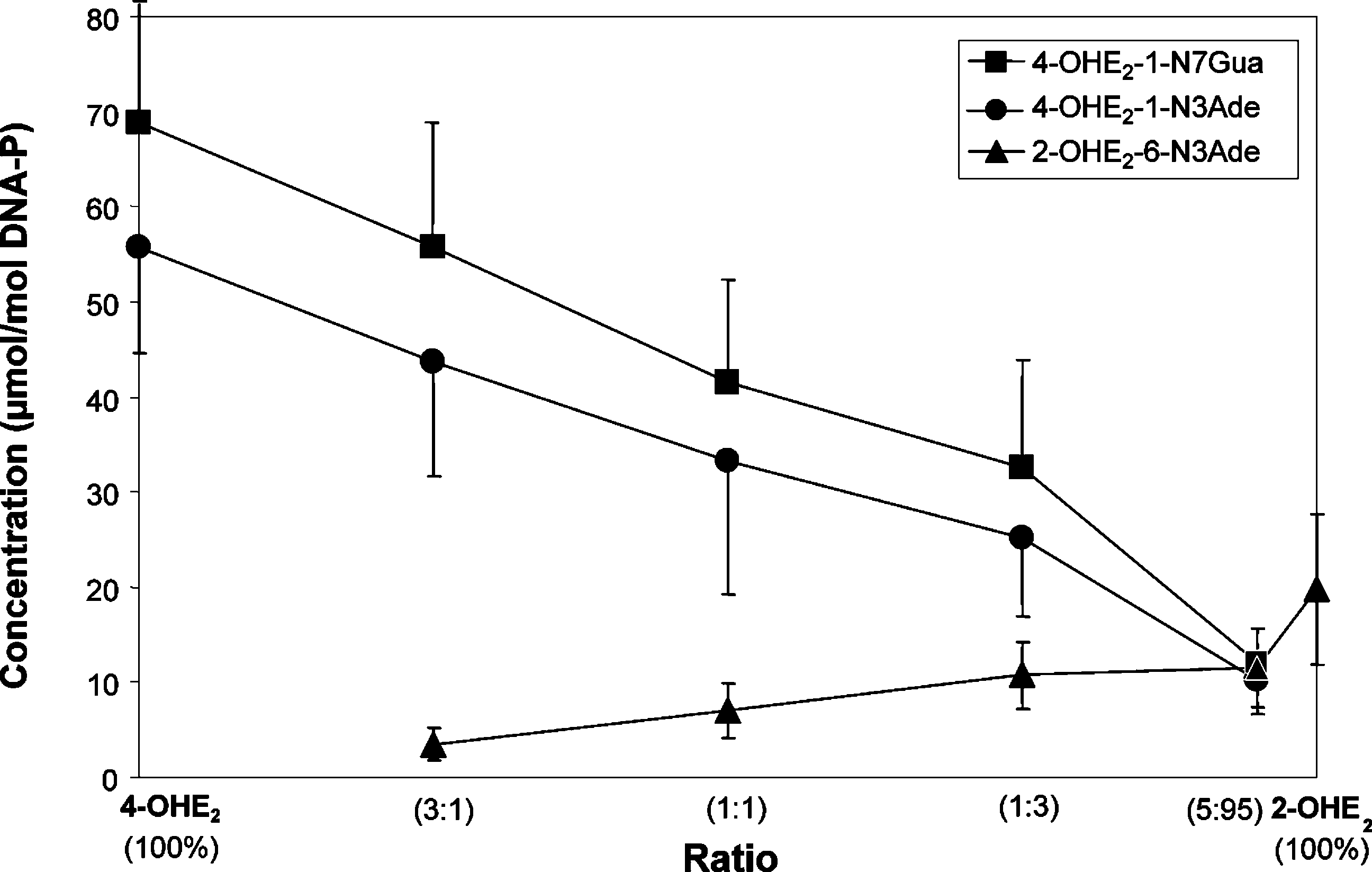
168 Chem. Res. Toxicol., Vol. 19, No. 1, 2006 Figure 3. Depurinating adducts formed by mixtures of E2-3,4-Q and E2-2,3-Q at different ratios after 10 h of reaction with DNA. The level of stable adducts formed in the mixtures ranged from 0.1 to 1% of total adducts. Mixtures containing 0.87 mM E2-3,4-Q + E2-2,3-Q and 3 mM DNA in 0.067 M sodium potassium phosphate, pH 7.0, were incubated at 37 °C. At the indicated times, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in the supernatant were analyzed.
estrogen moiety. This corroborates substitution at N-3 in the
C-17 hydroxy group and one triplet at 3.53 ppm for the C-17
proposed structure. The two signals at 6.80 and 6.30 ppm were
proton in the estrogen moiety were also present. The presence
assigned as the aromatic protons H-1 and H-4, respectively,
of a broad singlet at 7.53 ppm corresponding to one proton
based on 1D and 2D NMR studies, which ruled out the
suggests that the 6-NH is substituted in the proposed structure.
possibility of the substitution of Ade in the aromatic ring (at
The 1H NMR spectra of 2-OHE1-6-N3Ade and 2-OHE1-6-N6-
C-1 and C-4) of E2. Furthermore, the spectrum showed the
Ade are very similar to those of 2-OHE2-6-N3Ade and 2-OHE2-
doublet of one proton at 5.65 ppm, which correlated to a methine
6-N6Ade, except that the former two show no signals for the
group at 51.3 ppm in the HSQC experiment. These 1H and 13C
C-17 hydroxy group and C-17 proton. Thus, the structure of
chemical shifts suggested a substitution of the base at the C-6
2-OHE1-6-N3Ade and 2-OHE1-6-N6Ade is also established. All
position of E2. This is in accordance with the earlier observation
four compounds showed the desired exact [M + H]+ in their
that E1-2,3-Q undergoes tautomerization to its quinone methide
(26); thus, the value of 5.65 ppm was assigned to the H-6 of
Relative Reactivity of E2-3,4-Q and E2-2,3-Q with DNA.
E2. Furthermore, the resonance of this proton shows COSY
Different concentrations of E2-3,4-Q were reacted with 3 mM
correlation with aliphatic protons of the steroid skeleton at 1.76
DNA to examine the saturation level in the formation of
ppm, assigned as H-7 in COSY and TOCSY experiments. In
depurinating adducts (4-OHE2-1-N3Ade and 4-OHE2-1-N7Gua).
addition, H-7 showed correlations in the COSY spectrum with
The level of 0.87 mM E2-3,4-Q was found to be saturating for
two other protons, which give a mutual multiplet at 1.74-1.87
measurement of the rate of reaction between E
ppm. The broad signal at 7.20 ppm, exchangeable with D
corresponding to two protons in the 1H NMR spectrum, excludedthe possibility of attachment of the estrogen moiety at the NH
Incubations were conducted at 37 °C for 10 h because the
of Ade. This was further confirmed by an NOE experiment, in
N7Gua adduct depurinates slowly. When E2-3,4-Q was reacted
which irradiation of the signal at δ 6.30 (H-4) produced a strong
with DNA for 10 h, the adducts 4-OHE2-1-N3Ade and 4-OHE2-
effect on the signal at δ 7.47 (H-2, Ade) and vice versa. These
1-N7Gua were detected at approximately equal levels of 130-
extensive NMR studies unequivocally establish the structure of
140 µmol/mol DNA-P (data not shown).
the compound as 2-OHE2-6-N3Ade. Although the nucleophilic
The slow depurination of 4-OHE2-1-N7Gua is clearly seen
attack of the N-3 position of Ade can take place at either face
in Figure 2, in which equal amounts of E2-3,4-Q and E2-2,3-Q
at C-6 of the estrogen quinone methide, we obtained
were reacted with DNA and the depurinating adducts were
only one product, in which the attachment occurred at the R
analyzed at various time points. The binding of E2-3,4-Q to
face (from below the estrogen plane). Attack at the upper face
DNA and depurination of the N3Ade adduct was complete
appears to be hindered by the methyl group at C-13.
within 1 h, suggesting that depurination was instantaneous. In
The 1H NMR spectrum of 2-OHE2-6-N6Ade showed a broad
contrast, the depurination of the N7Gua adduct had a half-life
singlet integrated as two protons at 9.25 ppm for the two
of approximately 3 h at 37 °C and was complete in 10 h. This
hydroxy groups, exchangeable with D2O. Three singlets of one
slow loss of deoxyribose was previously observed in the reaction
proton each at 8.23, 8.07, and 7.73 ppm were assigned to the
of E2-3,4-Q with dG to form the N7Gua adduct (39). The adduct
H-8, H-2, and NH of the Ade moiety, respectively. Two singlets
2-OHE2-6-N3Ade was formed at a much lower level, 4 µmol/
at 6.67 and 6.55 ppm were assigned to the H-1 and H-4 protons
mol DNA-P, and depurination was also immediate (Figure 2).
of the E2 moiety, respectively; one broad singlet at 5.42 ppm
The level of stable adducts determined by the 32P-postlabeling
(one proton) for the H-6 of the E2 moiety strongly suggests
technique was <0.5% of the total adducts formed (data not
substitution at the C-6 position; one singlet at 4.40 ppm for the
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNAChem. Res. Toxicol., Vol. 19, No. 1, 2006 169 Figure 4. Depurinating adducts formed in the presence of tyrosinase by mixtures of 4-OHE2 and 2-OHE2 at different ratios after 10 h of reaction with DNA. The levels of stable adducts formed in the mixtures ranged from 0.1 to 0.7% of total adducts. Mixtures containing 0.87 mM 4-OHE2
+ 2-OHE2, 3 mM DNA, and 0.1 mg/mL tyrosinase in 0.067 M sodium potassium phosphate, pH 7.0, were incubated at 37 °C. At the indicatedtimes, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in the supernatant were analyzed. Figure 5. Depurinating adducts formed in the presence of LP by mixtures of 4-OHE2 and 2-OHE2 at different ratios after 10 h of reaction with DNA. The levels of stable adducts formed in the mixtures ranged from 0.2 to 0.8% of total adducts. Mixtures containing 0.87 mM 4-OHE +
2-OHE2, 3 mM DNA, and 0.1 mg/mL LP and 0.5 mM H2O2 in 0.067 M sodium potassium phosphate, pH 7.0, were incubated at 37 °C. At theindicated times, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in the supernatant were analyzed.
This study was further delineated by determining the level
(data not shown). Similar relative amounts of the three depu-
of the three depurinating adducts formed at different ratios of
rinating adducts were obtained when LP was used to activate
E2-3,4-Q and E2-2,3-Q (Figure 3). The overwhelming abundance
the catechol estrogens (Figure 5). The amounts of stable adducts
of the depurinating N7Gua and N3Ade adducts formed by E2-
formed in the mixtures were similar, ranging from 0.2 (4-OHE2)
3,4-Q is observed at all ratios of quinones. The levels of the
to 0.8% (2-OHE2) of total adducts (data not shown). With
three depurinating adducts were similar only with 5% E2-3,4-Q
activation by PHS, relatively lower amounts of adducts were
and 95% E2-2,3-Q. The level of stable adducts formed in these
formed from the 4-OHE2, as compared to activation by
mixtures ranged from 0.1% of total adducts with 100% E2-3,4-Q
tyrosinase or LP. The level of adducts from 2-OHE2 remained,
to 1% of total adducts with 100% E2-2,3-Q (data not shown).
instead, about the same (Figure 6). In this experiment, with 95%
Relative Reactivity of Enzyme-Activated 4-OHE2 and
2-OHE2 and 5% 4-OHE2, the levels of the two depurinating
2-OHE2 with DNA. When mixtures of 2-OHE2 and 4-OHE2
adducts from 4-OHE2 were about half the amount of 2-OHE2-
at different ratios were reacted with DNA in the presence of
6-N3Ade. The level of stable adducts was a little higher, and
tyrosinase, the depurinating adducts from 4-OHE2 were the most
they represented 0.5 (4-OHE2) to 4% (2-OHE2) of total adducts
abundant. The levels of the depurinating adducts were similar
with 95-100% 2-OHE2 (data not shown).
only with 95% 2-OHE2 and 5% 4-OHE2 present (Figure 4).
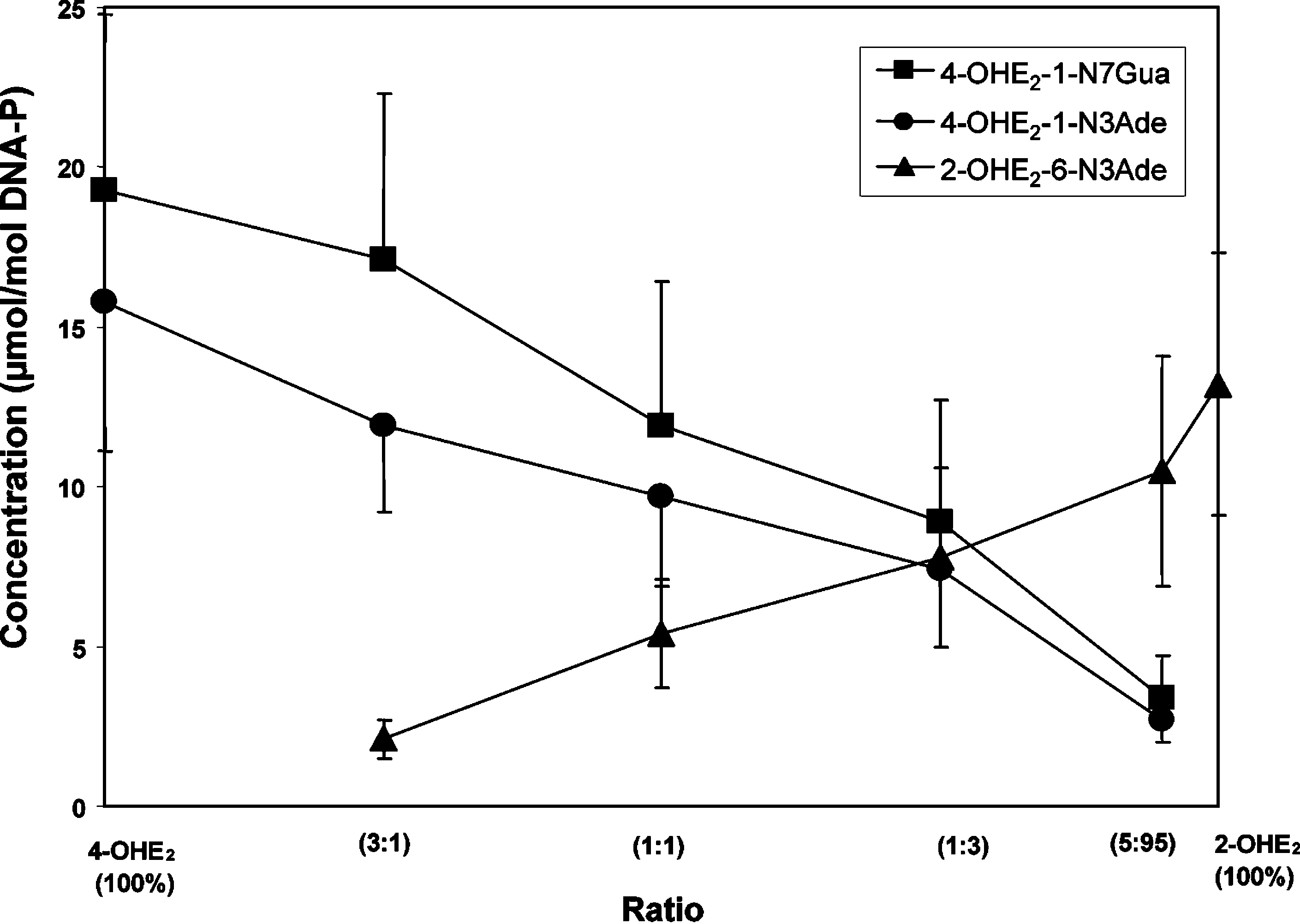
The activation of 4-OHE2 was relatively lower when MC-
The levels of stable adducts detected in the reaction mixtures
induced rat liver microsomes were used for activation (Figure
were approximately the same, which ranged from 0.1 (with
7). In fact, 100% 4-OHE2 yielded only 15-20 µmol/mol DNA-P
100% 4-OHE2) to 0.7% (with 100% 2-OHE2) of total adducts
of 4-OHE2-1-N7Gua and 4-OHE2-1-N3Ade. The 2-OHE2-6-
170 Chem. Res. Toxicol., Vol. 19, No. 1, 2006 Figure 6. Depurinating adducts formed in the presence of PHS by mixtures of 4-OHE2 and 2-OHE2 at different ratios after 10 h of reaction with DNA. The levels of stable adducts formed in the mixtures ranged from 0.5 to 4.1% of total adducts. Mixtures containing 0.87 mM 4-OHE +
2-OHE2, 3 mM DNA, 0.3 mg/mL methemoglobin, 5 mM arachidonic acid, and 40 units/mL PHS in 0.067 M sodium potassium phosphate, pH 7.0,were incubated at 37 °C. At the indicated times, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in the supernatantwere analyzed. Figure 7. Depurinating adducts formed in the presence of MC-induced rat liver microsomes by mixtures of 4-OHE2 and 2-OHE2 at different ratios after 10 h of reaction with DNA. The levels of stable adducts formed in the mixtures ranged from 0.3 to 11% of total adducts. Mixtures containing 0.87 mM 4-OHE +
2-OHE2, 3 mM DNA, 150 mM KCl, 5 mM MgCl2, 1 mg/mL microsomal protein, and 0.6 mM NADPH in 150 mM Tris-HCl,
pH 7.5, were incubated at 37 °C. At the indicated times, DNA was precipitated with 2 volumes of ethanol and the depurinating adducts in thesupernatant were analyzed.
N3Ade was obtained in relatively larger amounts. In fact, at a
the formation of stable adducts does not correlate with the
ratio of 75% 2-OHE2 to 25% 4-OHE2, similar amounts of the
carcinogenic potency of 2-OHE2 and 4-OHE2 (1, 20, 21).
three depurinating adducts were detected (Figure 7), and with95% 2-OHE2, the 2-OHE2-6-N3Ade was observed to be present
Conclusions
in about a 3-fold higher amount than the two adducts formedfrom 4-OHE2. With the microsomes, the amount of stable
The reaction of E2-3,4-Q with dG forms the depurinating
adducts ranged from 0.12 µmol/mol DNA-P with 100% 4-OHE2
adduct 4-OHE2-1-N7Gua (13, 16), whereas the reaction of E2-
to 1.57 µmol/mol DNA-P with 100% 2-OHE2, representing 0.3
3,4-Q or E2-2,3-Q with Ade forms 4-OHE2-1-N3Ade (16) or
(with 4-OHE2) to 11.4% (with 2-OHE2) of total adducts (data
2-OHE2-6-N3Ade, respectively. The reaction of E2-3,4-Q with
not shown). This 10-fold increase in the amount of stable
DNA yields 4-OHE2-1-N3Ade, which is rapidly depurinated,
adducts formed from 2-OHE2 as compared to 4-OHE2 is similar
and 4-OHE2-1-N7Gua, which is depurinated slowly, with a half-
to results previously obtained when E2-2,3-Q or E2-3,4-Q was
life of about 3 h (Figure 2). The final amounts of the two adducts
reacted with DNA (40). In the experiments reported here, the
are very similar (130-140 µmol/mol DNA-P). The reaction of
level of stable adducts formed from enzyme-activated 4-OHE2
E2-2,3-Q with DNA affords 2-OHE2-6-N3Ade, which depuri-
ranged from about the same as to approximately 10-fold less
nates immediately. The maximum amount of this adduct is 12
than the amount of stable adducts formed from 2-OHE2. Thus,
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNAChem. Res. Toxicol., Vol. 19, No. 1, 2006 171
When mixtures of E2-3,4-Q and E2-2,3-Q (3:1, 1:1, 1:3, and
References
5:95) react with DNA, the levels of 4-OHE2-1-N7Gua and
(1) Liehr, J. G., Fang, W. F., Sirbasku, D. A., and Ari-Ulubelen, A. (1986)
4-OHE2-1-N3Ade decrease with increasing amounts of E2-2,3-Q
Carcinogenicity of catechol estrogens in Syrian hamsters. J. Steroid
but are always much greater than the level of 2-OHE2-6-N3Ade
(2) Nandi, S. (1978) Role of hormones in mammary neoplasia. Cancer
by tyrosinase, LP, or PHS (Figures 4-6) in the presence of
(3) Li, J. J. (1993) Estrogen carcinogenesis in hamster tissues: Update.
DNA, various levels of the adducts are obtained, but the levels
Endocr. ReV. 14, 94-95.
of the N3Ade and N7Gua depurinating adducts are much greater
(4) Lang, R., and Redmann, U. (1979) Nonmutagenicity of some sex
hormones in the Ames salmonella/microsome mutagenicity test. Mutat.
N3Ade is higher than the levels of 4-OHE2-1-N3Ade and
(5) Lang, R., and Reimann, R. (1993) Studies for a genotoxic potential
4-OHE2-1-N7Gua only with PHS and 95% 2-OHE2. With rat
of some endogenous and exogenous sex steroids. I. Communication:
liver microsomes, relatively more depurinating adducts are
Examination for the induction of gene mutations using the Ames
Salmonella/microsome test and the HGPRT test in V79 cells. EnViron.
2, with equal amounts of all three depuri-
nating adducts being observed at 25% 4-OHE2 and 75% 2-OHE2
(6) Drevon, C., Piccoli, C., and Montesano, R. (1981) Mutagenicity assays
(Figure 7). The above data are essential for predicting the
of estrogenic hormones in mammalian cells. Mutat. Res. 89, 83-90.
relative amounts of depurinating adducts found in vivo when
(7) Furth, J. (1982) Hormones as etiological agents in neoplasia. In Cancer.A ComprehensiVe Treatise. 1. Etiology: Chemical and Physical
various ratios of 2-OHE2 and 4-OHE2 are formed during the
Carcinogenesis (Becker, F. F., Ed.) Chapter 4, pp 89-134, Cancer
(8) Li, J. J., and Li, S. A. (1990) Estrogen carcinogenesis in hamster
depurinating adducts of Ade and Gua is in line with the
tissues: A critical review. Endocr. ReV. 11, 524-531.
(9) Nandi, S., Guzman, R. C., and Yang, J. (1995) Hormones and
carcinogenic effect of 4-OHE2 in inducing kidney tumors in
mammary carcinogenesis in mice, rats and humans: A unifying
hamsters (1, 20) and uterine adenocarcinomas in CD-1 mice
hypothesis. Proc. Natl. Acad. Sci. U.S.A. 92, 3650-3657.
(21). The much lower reactivity of E
(10) Feigelson, H. S., and Henderson, B. E. (1996) Estrogens and breast
cancer. Carcinogenesis 17, 2279-2284.
2-OHE2-6-N3Ade also correlates with the lack of induction of
(11) Dickson, R. B., and Stancel, G. M. (2000) Estrogen receptor-mediated
kidney tumors by 2-OHE2 in hamsters (1, 20) and the much
processes in normal and cancer cells. In JNCI Monograph 27:
weaker ability to induce uterine adenocarcinomas in CD-1 mice
Estrogens as Endogenous Carcinogens in the Breast and Prostate
(21). Furthermore, in breast tissue from women with breast
(Cavalieri, E., and Rogan, E., Eds) pp 135-145, Oxford Press, NewYork.
carcinoma, 4-OHE1(E2) were 3.5 times more abundant than
(12) Hahn, W. C., and Weinberg, R. A. (2002) Rules for making tumor
2-OHE1(E2) and were four times higher than in breast tissue
cells. N. Engl. J. Med. 347, 1593-1603.
from women without breast cancer (41, 42).
(13) Cavalieri, E. L., Stack, D. E., Devanesan, P. D., Todorovic, R.,
Dwivedy, I., Higginbotham, S., Johansson, S. L., Patil, K. D., Gross,
When cultured human breast epithelial (MCF-10F) cells,
M. L., Gooden, J. K., Ramanathan, R., Cerny, R. L., and Rogan, E.
which are estrogen receptor-R negative, are treated with
G. (1997) Molecular origin of cancer: Catechol estrogen-3,4-quinones
as endogenous tumor initiators. Proc. Natl. Acad. Sci. U.S.A. 99,
An approximately million times higher concentration of 2-OHE
(14) Cavalieri, E., Frenkel, K., Liehr, J. G., Rogan, E., and Roy, D. (2000)
is needed to transform these cells (22, 23). Transformation
Estrogens as endogenous genotoxic agents:
occurs even in the presence of the antiestrogen ICI-182,780 (24),
indicating that transformation does not proceed through estrogen
Carcinogens in the Breast and Prostate (Cavalieri, E., and Rogan,E., Eds.) pp 75-93, Oxford Press, New York.
receptor-mediated events (25). In fact, specific mutations are
(15) Cavalieri, E., Rogan, E., and Chakravarti, D. (2004) The role of
induced by treatment of MCF-10F cells with 4-OHE2 (25).
endogenous catechol quinones in the initiation of cancer and neuro-
degenerative diseases. In Methods in Enzymology, Quinones and
2 and E2-3,4-Q have been shown to induce mutations
Quinone Enzymes, Part B (Sies, H., and Packer, L., Eds) Vol. 382,
in several animal models. Topical treatment of SENCAR mouse
pp 293-319, Elsevier, Duesseldorf, Germany.
skin with E2-3,4-Q induces A f G mutations in the Harvey-
(16) Li, K.-M., Todorovic, R., Devanesan, P., Higginbotham, S., Ko¨feler,
ras reporter gene within 12 h (17), and similar mutations are
H., Ramanathan, R., Gross, M. L., Rogan, E. G., and Cavalieri, E. L.
observed in ACI rat mammary glands within 12 h of intramam-
(2004) Metabolism and DNA binding studies of 4-hydroxyestradioland estradiol-3,4-quinone in vitro and in female ACI rat mammary
millary treatment of the mammary glands with E2-3,4-Q (18).
gland in vivo. Carcinogenesis 25, 289-297.
Recently, 4-OHE2 was found to be mutagenic in the mammary
(17) Chakravarti, D., Mailander, P., Li, K.-M., Higginbotham, S., Zhang,
gland of female Big Blue rats after 20 weeks of treatment with
H., Gross, M. L., Cavalieri, E., and Rogan, E. (2001) Evidence that aburst of DNA depurination in SENCAR mouse skin induces error-
implanted estrogen. Much higher fractions of A f G mutations
prone repair and forms mutations in the H-ras gene. Oncogene 20,
were found in the mammary glands of the 4-OHE2-treated rats
than controls (J. Guttenplan, private communication). Addition-
(18) Chakravarti, D., Mailander, P. C., Higginbotham, S., Cavalieri, E. L.,
and Rogan, E. G. (2003) The catechol estrogen-3,4-quinone metabolites
2, but not 2-OHE2, was found to be mutagenic in
induce mutations in the mammary gland of ACI rats. Proc. Am. Assoc.
Big Blue rat 2 embryonic cells, and the mutants contained
significantly higher fractions of A f G mutations than those
(19) Zhao, Z., Kosinska, W., Khmelnitsky, M., Cavalieri, E. L., Rogan, E.
G., Chakravarti, D., Sacks, P., and Guttenplan, J. B. (2005) Mutagenic
2-3,4-Q was also mutagenic. In summary, there
activity of 4-hydroxyestradiol, but not 2-hydroxyestradiol, in BB2 rat
is a general correlation between the depurinating adducts formed
embryonic cells, and the mutational spectrum of 4-hydroxyestradiol.
by oxidation of 4-OHE2 to E2-3,4-Q and the mutagenicity in
Chem. Res. Toxicol., in press.
vitro and in vivo, cell transformation, and carcinogenicity by
(20) Li, J. J., and Li, S. A. (1987) Estrogen carcinogenesis in Syrian hamster
tissue: Role of metabolism. Fed. Proc. 46, 1858-1863.
(21) Newbold, R. R., and Liehr, J. G. (2000) Induction of uterine
adenocarcinoma in CD-1 mice by catechol estrogens. Cancer Res. 60,
Acknowledgment. This research was supported by Grant
P01 CA49210 from the National Cancer Institute and Grant
(22) Russo, J., Lareef, M. H., Tahin, Q., Hu, Y.-F., Slater, C., Ao, X., and
DAMD17-03-10229 from the Department of Defense Breast
Russo, I. H. (2002) 17 -Estradiol is carcinogenic in human breastepithelial cells. J. Steroid Biochem. Mol. Biol. 80, 149-162.
Cancer Research Program. Core support at the Eppley Institute
(23) Russo, J., Lareef, M. H., Balogh, G., Guo, S., and Russo, I. H. (2003)
was provided by Grant P30 CA36727 from the National Cancer
Estrogen and its metabolites are carcinogenic agents in human breast
epithelial cells. J. Steroid Biochem. Mol. Biol. 87, 1-25. 172 Chem. Res. Toxicol., Vol. 19, No. 1, 2006
(24) Lareef, M. H., Garber, J., Russo, P. A., Russo, I. H., Heulings, R.,
determination of the adducts formed by electrochemical oxidation of
and Russo, J. (2005) The estrogen antagonist ICI-182-780 does not
1,2,3,4-tetrahydro-7,12-dimethylbenz[a]anthracene in the presence of
inhibit the transformation phenotypes induced by 17- -estradiol and
deoxyribonucleosides or adenine. Chem. Res. Toxicol. 9, 1264-1277.
4-OH estradiol in human breast epithelial cells. Int. J. Oncol. 26, 423-
(35) Hanson, A. A., Rogan, E. G., and Cavalieri, E. L. (1998) Synthesis
of adducts formed by iodine oxidation of aromatic hydrocarbons in
(25) Russo, I. H., Fernandez, S. V., and Russo, J. (2005) Estradiol and its
the presence of deoxyribonucleosides and nucleobases. Chem. Res.
metabolites 4-hydroxyestradiol and 2-hydroxyestradiol induce muta-
tions in human breast epithelial cells. Int. J. Cancer, in press.
(36) Li, K.-M., Byun, J., Gross, M. L., Zamzow, D., Jankowiak, R., Rogan,
(26) Stack, D., Byun, J., Gross, M. L., Rogan, E. G., and Cavalieri, E.
E. G., and Cavalieri, E. L. (1999) Synthesis and structure determination
(1996) Molecular characteristics of catechol estrogen quinones in
of the adducts formed by electrochemical oxidation of dibenzo[a,l]-
reactions with deoxyribonucleosides. Chem. Res. Toxicol. 9, 851-
pyrene in the presence of adenine. Chem. Res. Toxicol. 12, 749-757.
(37) Chen, L., Devanesan, P. D., Higginbotham, S., Ariese, F., Jankowiak,
(27) NIH Guidelines for the Laboratory Use of Chemical Carcinogens
R., Small, G. J., Rogan, E. G., and Cavalieri, E. L. (1996) Expanded
(1981) NIH Publication No. 81-2385, U.S. Government Printing
analysis of benzo[a]pyrene-DNA adducts formed in vitro and in mouse
skin: Their significance in tumor initiation. Chem. Res. Toxicol. 9,
(28) Saeed, M., Zahid, M., Rogan, E., and Cavalieri, E. (2005) Synthesis
of the catechols of natural and synthetic estrogens by using 2-iodoxy-
(38) Cavalieri, E. L., Rogan, E. G., Li, K.-M., Todorovic, R., Ariese, F.,
benzoic acid (IBX) as the oxidizing agent. Steroids 70, 173-178.
Jankowiak, R., Grubor, N., and Small, G. J. (2005) Identification and
(29) Wong, A. K. L., Cavalieri, E., and Rogan, E. (1986) Dependence of
quantification of the depurinating DNA adducts formed in mouse skin
benzo[a]pyrene metabolic profile on the concentration of cumene
treated with dibenzo[a,l]pyrene (DB[a,l]P) or its metabolites and in
hydroperoxide with uninduced and induced rat liver microsomes.
rat mammary gland treated with DB[a,l]P. Chem. Res. Toxicol. 18,
Biochem. Pharmacol. 35, 1583-1588.
(30) Mancera, O., Rosenkranz, G., and Sondheimer, F. (1953) Steroids.
(39) Saeed, M., Zahid, M., Gunselman, S. J., Rogan, E., and Cavalieri, E.
Part XLVI. Synthesis of 11 -hydroxytestosterone and 11-keto test-
(2005) Slow loss of deoxyribose from the N7deoxyguanosine adducts
osterone. J. Chem. Soc. Abstract 2189-2191.
of estradiol-3,4-quinone and hexestrol-3′,4′-quinone. Implications for
(31) Bodell, W. J., Devanesan, P. D., Rogan, E. G., and Cavalieri, E. L.
mutagenic activity. Steroids 70, 29-35.
(1989) 32P-Postlabeling analysis of benzo[a]pyrene-DNA adducts
(40) Dwivedy, I., Devanesan, P., Cremonesi, P., Rogan, E., and Cavalieri,
formed in vitro and in vivo. Chem. Res. Toxicol. 2, 312-315.
E. (1992) Synthesis and characterization of estrogen 2,3- and 3,4-
(32) Saeed, M., Gunselman, S. J., Higginbotham, S., Rogan, E., and
quinones. Comparison of DNA adducts formed by the quinones versus
Cavalieri, E. (2005) Formation of the depurinating N3adenine and
horseradish peroxidase-activated catechol estrogens. Chem. Res.
N7guanine adducts by reaction of DNA with hexestrol-3′-4′-quinone
or enzyme-activated 3′-hydroxyhexestrol. Implications for a unifying
(41) Liehr, J. G., and Ricci, M. J. (1996) 4-Hydroxylation of estrogens as
mechanism of tumor initiation by natural and synthetic estrogens.
markers of human mammary tumors. Proc. Natl. Acad. Sci. U.S.A.
(33) RamaKrishna, N. V., Padmavathi, N. S., Cavalieri, E. L., Rogan, E.
(42) Rogan, E. G., Badawi, A. F., Devanesan, P. D., Meza, J. L., Edney,
G., Cerny, R. L., and Gross, M. L. (1993) Synthesis and structure
J. A., West, W. W., Higginbotham, S. M., and Cavalieri, E. L. (2003)
determination of the adducts formed by electrochemical oxidation of
Relative imbalances in estrogen metabolism and conjugation in breast
the potent carcinogen dibenzo[a,l]pyrene in the presence of nucleo-
tissue of women with carcinoma: Potential biomarkers of susceptibility
sides. Chem. Res. Toxicol. 6, 554-560.
to cancer. Carcinogenesis 24, 697-702.
(34) Mulder, P. P., Chen, L., Sekhar, B. C., George, M., Gross, M. L.,
Rogan, E. G., and Cavalieri, E. L. (1996) Synthesis and structure
Nutrition and Cancer Brown Kelp Modulates Endocrine Hormones in Female Sprague-Dawley Rats and in Human Luteinized Granulosa Cells1 Christine F. Skibola,*2 John D. Curry,*3 Catherine VandeVoort,† Alan Conley,** andMartyn T. Smith* *School of Public Health, University of California, Berkeley, California; and †California National PrimateResearch Center and **Department of Population H
SLEEP APNEA SYNDROMES The mysteries of sleep have intrigued man for centuries, it has only been within last three decades that we have begun to understand physiology and physiopatology of this state that occupies a considerable portion of our lives. It was developped a better understanding of specific sleep disorders and their realationship to common medical problems seen during wakefulnes
 ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 165
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 165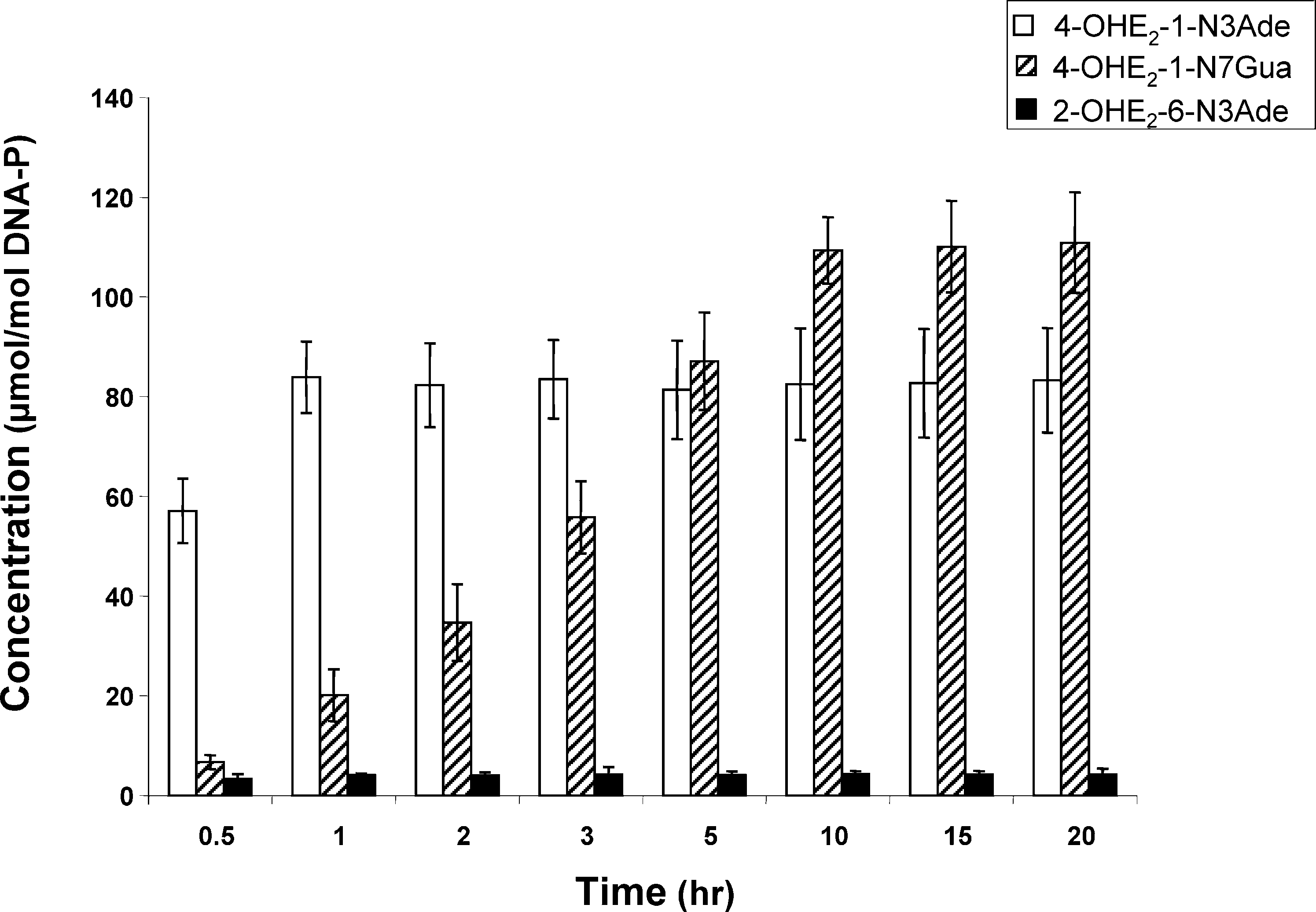 ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 167
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 167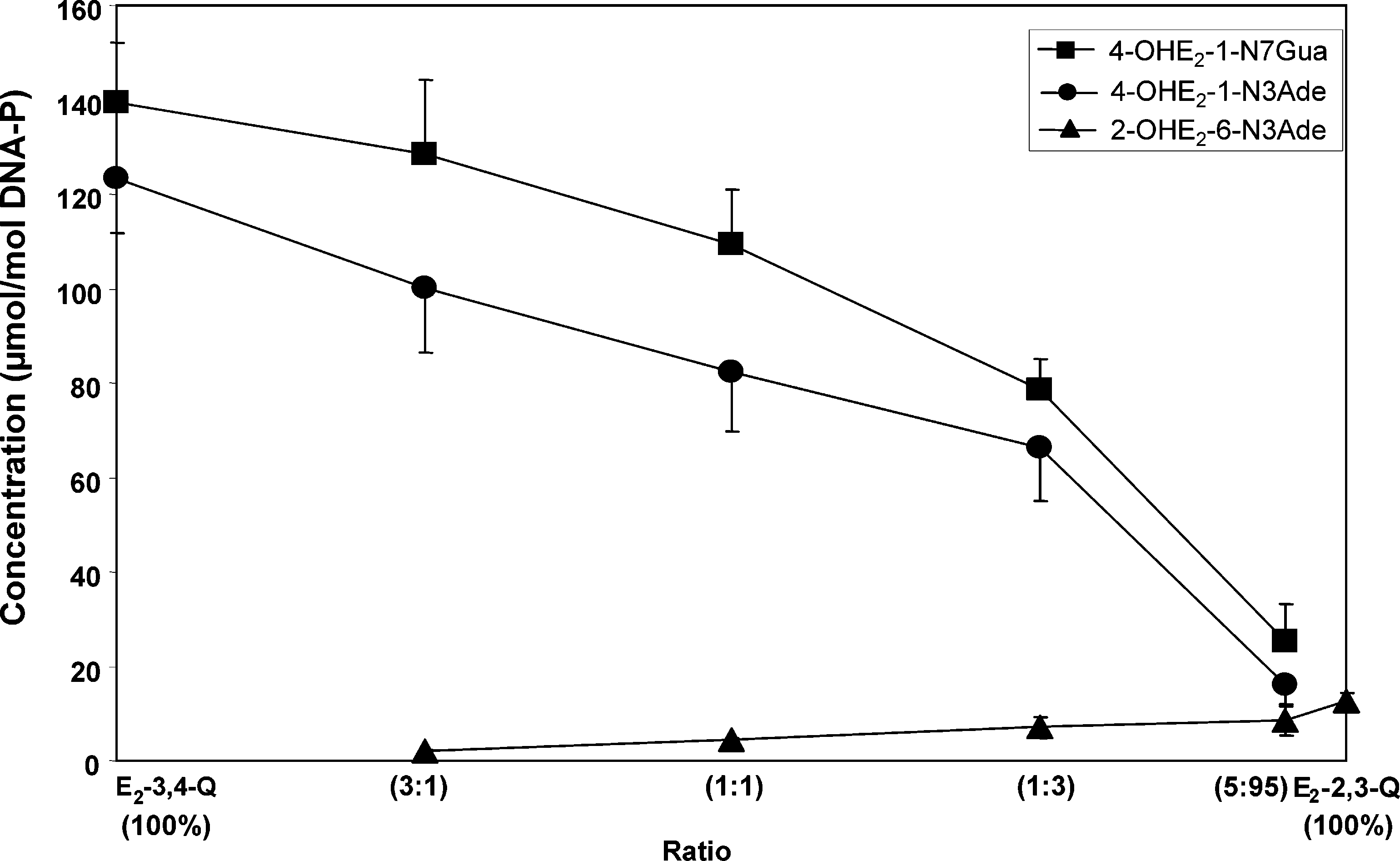 168 Chem. Res. Toxicol., Vol. 19, No. 1, 2006
168 Chem. Res. Toxicol., Vol. 19, No. 1, 2006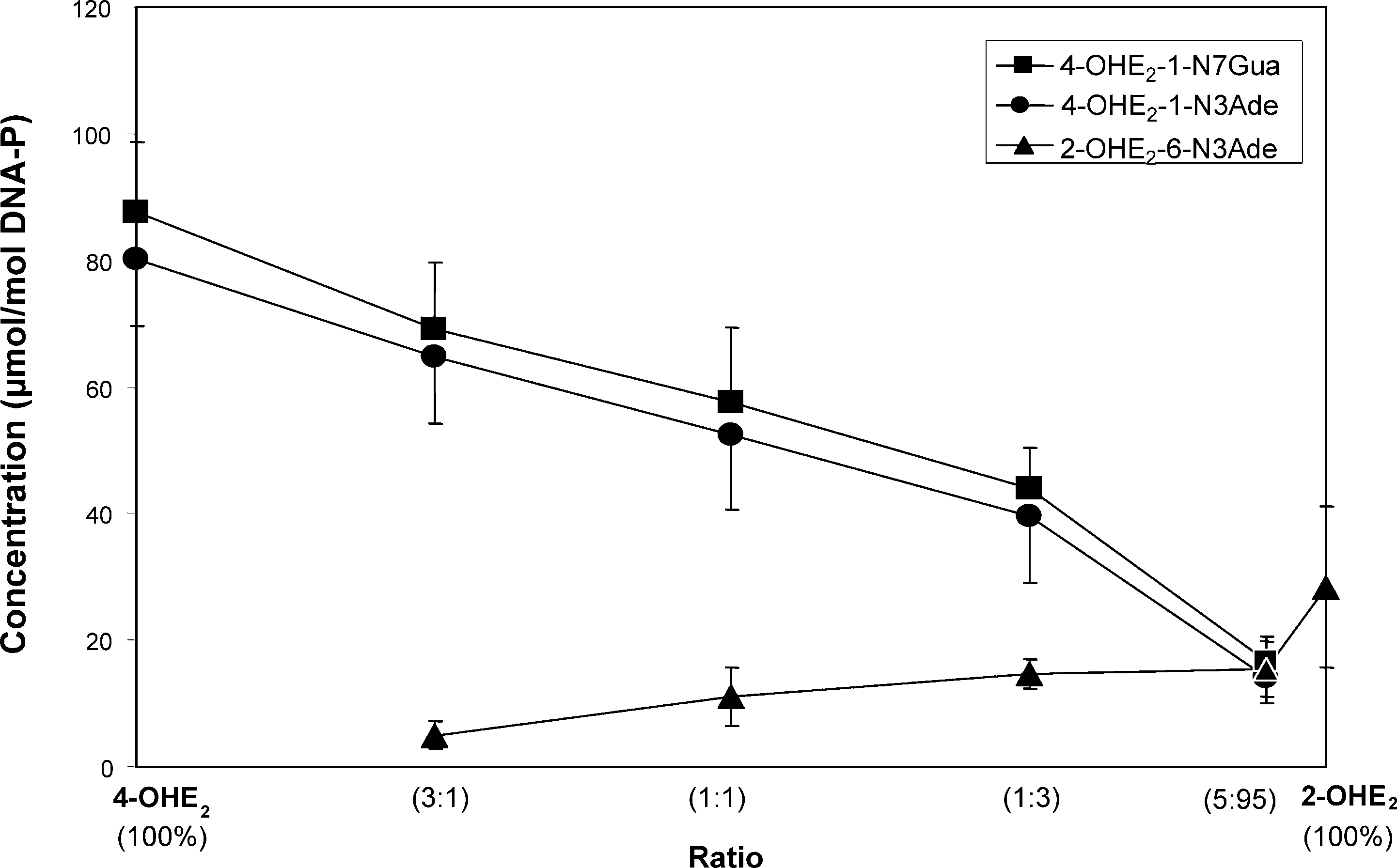
 ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 169
ReactiVity of E2-3,4-Q Vs E2-2,3-Q with DNA
Chem. Res. Toxicol., Vol. 19, No. 1, 2006 169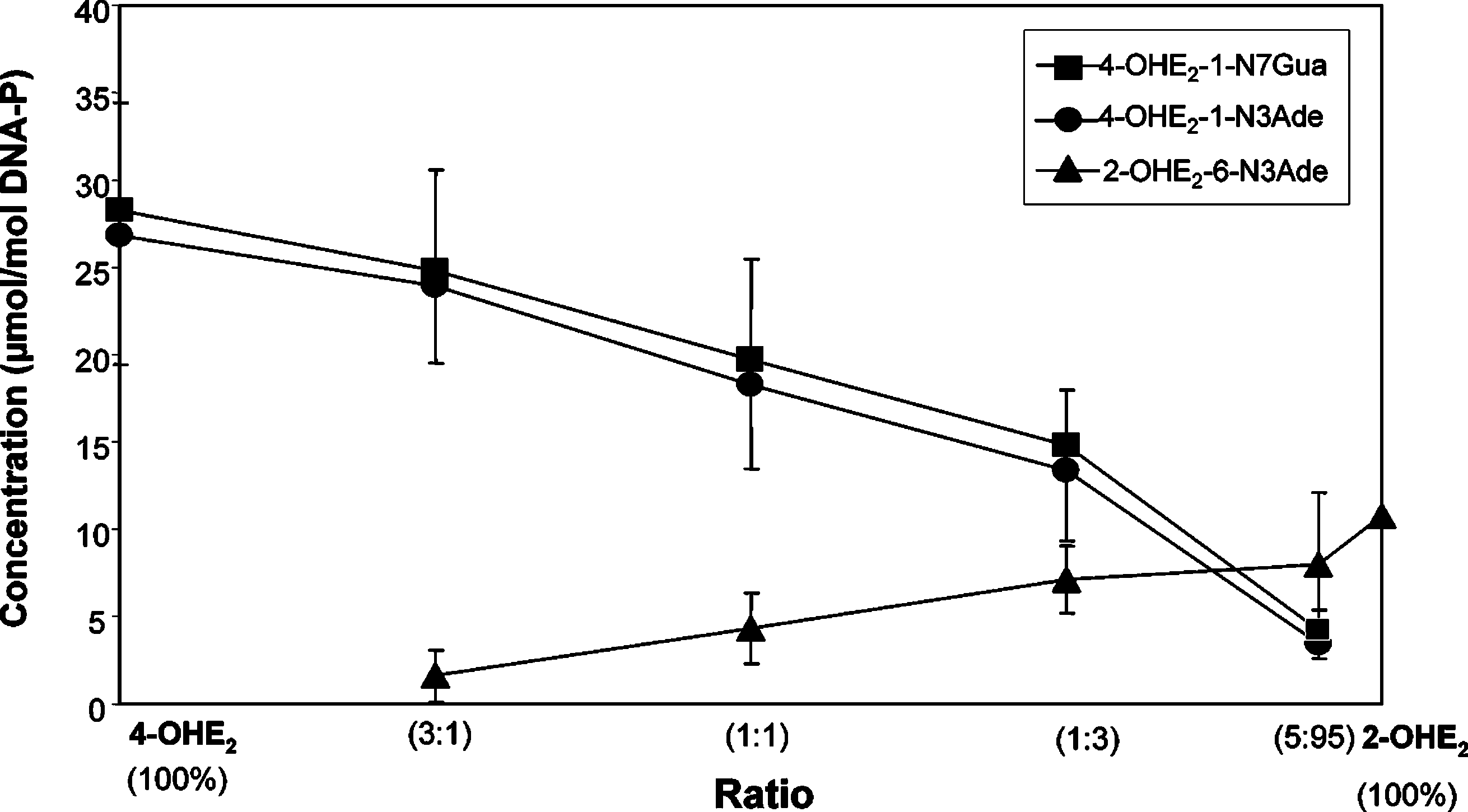
 170 Chem. Res. Toxicol., Vol. 19, No. 1, 2006
170 Chem. Res. Toxicol., Vol. 19, No. 1, 2006