Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Nbmu.ucsd.edu
Neuropsychopharmacology (2004) 29, 731–738& 2004 Nature Publishing Group
Reversal of Sensorimotor Gating Deficits in Brattleboro Ratsby Acute Administration of Clozapine and a NeurotensinAgonist, but not Haloperidol: a Potential Predictive Model forNovel Antipsychotic Effects
David Feifel*,1, Gilia Melendez1 and Paul D Shilling1
Department of Psychiatry, University of California, San Diego, San Diego, CA, USA
Prepulse inhibition (PPI) of acoustic startle is decreased in unmedicated schizophrenia patients and similar deficits can be induced in rats
through pharmacological, environmental, or neuroanatomical manipulations. Recently, we reported that Brattleboro (BB) rats, a Long
Evans (LE) strain with a single gene mutation, have inherent deficits in PPI homologous to those observed in schizophrenia patients. We
also reported that PPI deficits in BB rats could be reversed by chronic but not acute administration of 0.5 mg/kg haloperidol. No other
dose or drug was tested in that experiment. In this study, we tested the effects of acute subcutaneous administration of several doses of
haloperidol as well as the second-generation antipsychotic, clozapine, and the putative novel antipsychotic, PD149163, a neurotensin
mimetic that crosses the blood–brain barrier. Consistent with our previous report, BB rats exhibited PPI deficits compared to LE rats and
none of the doses of haloperidol produced a significant effect on this PPI deficit. In contrast, 10 and 15 mg/kg of clozapine and all the
doses of PD149163 tested reversed the PPI deficits in BB rats. In addition, haloperidol, but not clozapine or PD149163 produced
significant catalepsy in BB rats, supporting the notion that PD149163 has a profile consistent with atypical antipsychotics and providing
support for the predictive validity of the PPI results. These results further strengthen the notion that the BB rat is a useful predictive
model of antipsychotic efficacy and suggest that this model may differentiate between antipsychotics belonging to different therapeutic
categories, for example, first- and second-generation agents.
Neuropsychopharmacology (2004) 29, 731–738, advance online publication, 4 February 2004; doi:10.1038/sj.npp.1300378
Keywords: antipsychotics; Brattleboro rats; prepulse inhibition; clozapine; neurotensin; schizophrenia
families including dopamine agonists such as amphetamineand apomorphine (Mansbach et al, 1988), serotonin
Prepulse inhibition (PPI) of the acoustic startle reflex is the
agonists such as DOI (Sipes and Geyer, 1994), and
reduction in the startle response when the startle-eliciting
noncompetitive NMDA antagonists such as phencyclidine
stimulus is immediately preceded by a weak stimulus
(PCP) and dizocilpine (MK801) (Mansbach and Geyer,
(Swerdlow and Geyer, 1998). PPI, an operational measure of
1989). Antipsychotics can reverse this disruption, making
sensorimotor gating, is deficient in schizophrenia patients,
PPI the basis of a predictive model for antipsychotic drugs
a phenomenon thought to reflect a disruption in cortico-
(Geyer et al, 2001). PPI deficits produced by dopamine
striatal-pallidal-pontine circuits involved in preconscious
agonists can be reversed by first-generation or ‘typical’ as
processing of environmental stimuli (Geyer et al, 2001). PPI
well as second-generation or ‘atypical’ antipsychotics;
deficits analogous to those seen in schizophrenia (Braff and
therefore, this paradigm is not useful in distinguishing
Geyer, 1990) can be induced in rats by administering
between these two categories of antipsychotics. In contrast,
psychomimetic drugs of several different pharmacological
PPI deficits produced by serotonin agonists and NMDAantagonists tend to be preferentially reversed by atypical
*Correspondence: Dr D Feifel, Department of Psychiatry, University of
antipsychotics (Geyer et al, 2001). Therefore, the reversal of
California, San Diego, 200 West Arbor Drive, San Diego, CA 92103-
serotonin agonist and NMDA antagonist-induced PPI
8218, USA, Tel: þ 1 619 543 2485; Fax: þ 1 619 543 3738,
disruption may be predictive of atypical antipsychotic drug
Received 08 July 2003; revised 30 October 2003; accepted 04December 2003
Paradigms requiring drugs to induce PPI deficits have
Online publication: 16 December 2003 at http://www.acnp.org/
certain inherent limitations as models of sensorimotor
gating deficits in humans and predictive models of
Reversal of sensorimotor gating deficits in BB rats
antipsychotic potential. Therefore, there is significant
effects on PPI are stable across the circadian cycle (Weiss
interest in development of nonpharmacological paradigms
et al, 1999). The rats were tested in startle chambers to
of PPI deficits. Two of the most studied examples of
characterize their baseline PPI and startle. Animals
nonpharmacological paradigms of PPI deficits are social
were assigned, based on their baseline PPI, to one of four
isolation-rearing (Geyer et al, 1993) and neonatal hippo-
groups matched so as to achieve comparable average PPI
campal lesions (Lipska et al, 1995). PPI deficits produced by
across groups. Drug treatment began 3 days after baseline
social isolation rearing are reversed by both typical and
atypical antipsychotics (Cilia et al, 2001; Le Pen and
In one experiment 50 LE rats and 50 BB rats were
Moreau, 2002). Preliminary findings suggest that PPI
administered subcutaneous (s.c.) injections of either 0
deficits produced by neonatal hippocampal lesions may be
(vehicle), 0.1, 0.5, or 1 mg/kg of haloperidol (UCSD Medical
reversed by atypical but not typical antipsychotics (Le Pen
Center, San Diego, CA). In another experiment 51 LE and 49
BB rats were administered s.c. injection of either 0 (vehicle),
Brattleboro (BB) rats are Long Evans (LE) rats with a
5, 10, or 15 mg/kg clozapine (Sigma Chemicals, St Louis,
single base pair mutation that results in the inability to
MO). In a third study, 43 LE rats and 33 BB rats were
properly synthesize the neurotransmitter and neurohor-
administered s.c. injection of either 0 (vehicle), 0.5, 1, or
mone vasopressin. BB rats have many behavioral and
2 mg/kg PD149163 (LYS(CH2NH)LYS-PRO-TRP-tLE-LEU-
cognitive abnormalities, including deficits in memory
OEt) (SRI International, NIMH chemical synthesis pro-
(Laycock et al, 1983), emotion (Williams et al, 1985), social
gram). Doses were selected based on their demonstrated
recognition (Engelmann and Landgraf, 1994), motivation,
ability to reverse PPI deficits in other reports (Feifel et al,
and attention (Williams et al, 1983). They have abnormal-
1999; Geyer et al, 2001). Vehicle for haloperidol was distilled
ities in brain systems including dopamine and serotonin,
water and the volume injected was 1 ml/kg. Vehicle for
neurotransmitters implicated in schizophrenia (Feenstra
clozapine and PD149163 was 0.1 N HCl and half volume
et al, 1990). BB rats also have deficits in PPI compared with
0.9% saline brought to pH 5–6 with a few drops of 1 N
their wild-type, LE counterparts. We previously showed that
NaOH. The volume injected for clozapine was 1.5 ml/kg and
acute treatment with a single dose (0.5 mg/kg) of haloper-
for PD149163 was 1 ml/kg. Each treatment group had a
idol did not reverse the PPI deficits in BB rats. In contrast,
chronic treatment with that dose of haloperidol reverses the
Animals were tested in startle chambers (San Diego
deficits, suggesting that the BB rat may be a very useful
Instruments, San Diego, CA) 20 min after drug administra-
genetic model of sensorimotor gating deficits associated
tion. Once placed in startle chambers, each rat had a 5-min
with schizophrenia that models the therapeutic time course
acclimation period. A 65-dB background noise was
of antipsychotic drugs (Feifel and Priebe, 2001). Since only
continuously present throughout the session. The acclima-
one dose of haloperidol was used in that study, it is possible
tion was followed by a 15 min PPI test session during which
that acute administration of higher or lower doses of
rats were presented with 40 ms 120 dB startle pulses without
haloperidol may reverse the PPI deficits observed in BB rats.
a prepulse, or pulses preceded 100 ms by a prepulse of
Furthermore, it is not known whether haloperidol’s inability
either 4, 8, or 12 dB above background. These four types of
to reverse PPI deficits in BB rats after acute administration
active stimuli were presented in addition to a neutral (no
extends to other established or putative antipsychotic drugs.
sound) stimuli condition in pseudorandom order with an
Therefore, we performed dose response experiments to
average of 15 s between stimuli types.
determine if haloperidol, and the atypical antipsychotic,
A startle response was recorded for all stimuli presenta-
clozapine and the putative antipsychotic, PD149163, a
tions. PPI for each animal was calculated as a percentage of
neurotensin mimetic that has been shown in previous
the pulse-alone startle magnitude using the following
studies to display antipsychotic-like properties (Feifel et al,
formula: (1À(startle magnitude after prepulse-pulse pair/
1999), would attenuate BB PPI deficits. Furthermore, we
startle magnitude after pulse only) Â 100). Exploratory
tested the catalepsy effects of all three drugs in order to
analysis of the data was conducted and indicated that PPI
determine whether PD149163 resembles clozapine or
deficits in BB rats were consistently more robust in the first
haloperidol in this respect, and thus to facilitate interpreta-
half of the startle sessions. Therefore, PPI data from this
tion of the predictive validity of the PPI results.
first block of stimuli were subjected to the further statisticalanalysis. To compare treatments groups, PPI data wassubjected to a three-way ANOVA in which prepulse
intensity was a within-subject factor and strain andtreatment (ie drug dose) were between-subject factors. As
All experimental procedures were conducted in accordance
expected, percent PPI was inversely related to prepulse
with the University of California, San Diego guidelines for
intensity in all three experiments (main effect of prepulse
animal care and experimentation. In total, 132 male BB rats
intensity) and this is a well-established relationship.
and 144 LE rats (170–350 g at testing, Harlan Laboratories,
However, there was no significant two- or three- way
San Diego) were housed in groups of two or three in clear
interaction of prepulse intensity with any of the drugs
plastic chambers in a climate controlled room under a 12/
tested. Therefore, this term was dropped from the
12 h light/dark schedule (lights on/off – 0700/1900). They
model and the analysis reported is based upon a reduced
were allowed free access to food and water for the extent of
model examining averaged PPI from all prepulse inten-
the study. Behavioral testing was performed 7 days after
arrival, during the light phase of the rats’ circadian
Dunnett’s one-tailed test to test the following specific
illumination schedule as startle magnitude, PPI, and drug
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
1. PPI in untreated BB rats is significantly lower than in
2. Treatment with the test drugs facilitates PPI in BB rats
3. Treatment of BB rats with the test drugs restores their
Data of the acoustic startle response (ASR) to the startle
stimuli not preceded by any prepulse were subjected to
analysis using a similar two factor ANOVA.
In a separate study to compare the catalepsy effects of
each drug, 48 drug-naı¨ve BB rats were given one of the
following SC treatments: saline, 1 mg/kg haloperidol, 10 mg/kg clozapine, 15 mg/kg clozapine, 1 mg/kg PD149163 or
2 mg/kg PD149163 (n ¼ 8 for all groups). Doses selectedwere those that produced the greatest effect in the PPI
studies. At 30 minutes after s.c. injections, animals weretested using a method described by several other authors(Stanley and Glick, 1976; Costall et al, 1978; Wadenberg,
1996). This method involved placing the forepaws of each
rat in an extended position over a pencil that was suspended
horizontally 9 cm above the lab bench. The time spent inthis position before the animal moved or corrected itselfwas considered a measure of catalepsy.
Figure 1 (top) illustrates PPI results in the haloperidol
(F(1,92) ¼ 23.3, Po0.001) with LE rats exhibiting higher
PPI than BB rats. There was no significant main effect ofhaloperidol nor was there a significant haloperidol  strain
PPI (top) and ASR (bottom) 7SEM in rats treated with
haloperidol. Significantly different from vehicle-treated LE rats represented
interaction. PPI exhibited by vehicle-treated LE rats was
by *(Po0.05) and **(Po0.01). Significantly different from vehicle-treated
significantly higher (Po0.05) than BB rats for all doses of
haloperidol. None of the doses of haloperidol significantlyincreased PPI in either LE or BB rats.
Figure 1 (bottom) illustrates the ASR results in the
between BB and LE rats. Clozapine appeared to reduce ASR
haloperidol experiment. Analysis of the ASR data indicates
in both rat strains in a dose-dependent fashion, but this
that BB rats had significantly higher ASR (F(1,92) ¼ 31.6,
effect was slightly above the statistical cutoff for significance
Po0.001), and that haloperidol significantly decreased the
(P ¼ 0.051). There was no significant clozapine  strain
ASR in both strains of rats (F(3,92) ¼ 13.6, Po0.001). There
was no significant strain  haloperidol interaction.
Figure 3 (top) illustrates the PPI results in the PD149163
Figure 2 (top) illustrates the PPI results in the clozapine
experiment. There was a significant effect of strain
experiment. There was a significant main effect of strain
((F(1,68) ¼ 6.28, Po0.05), a significant main effect of
(F(1,92) ¼ 14.7, Po0.001) and a strain  clozapine interac-
PD149163 (F(3,68) ¼ 4.75, Po0.01), and a significant
tion (F(3,92) ¼ 3.4, Po0.05), but not a main clozapine
strain  PD149163 interaction (F(3,68) ¼ 4.221, Po0.01).
effect. LE rats treated with vehicle exhibited PPI that was
PPI in vehicle-treated LE rats was significantly higher than
significantly higher than BB rats treated with vehicle
in vehicle-treated BB rats (Po0.01), but not significantly
(Po0.01), but not significantly different than BB rats
different from BB rats treated with any of the doses of
treated with any of the doses of clozapine tested.
PD149163. BB rats treated with 1 and 3 mg/kg PD149163
Furthermore, PPI in BB rats treated with vehicle was
had significantly higher PPI (Po0.01 and Po0.05, respec-
significantly lower than PPI in BB rats treated with 10 mg/kg
tively) than PPI exhibited by vehicle-treated BB rats. In
(Po0.01) and 15 mg/kg dose (Po0.05) of clozapine. In
contrast, no dose of PD149163 had a significant effect on
contrast, no dose of clozapine increased PPI in LE rats.
PPI in LE rats, although there appeared to be a tendency for
Figure 2 (bottom) illustrates the ASR results in the
PD149163 to dose-dependently increase PPI in LE rats.
clozapine experiment. Analysis of the ASR data indicates
Figure 3 (bottom) illustrates the ASR data for the
that there was no significant difference in startle magnitude
PD149163 experiment. There was not a significant differ-
Reversal of sensorimotor gating deficits in BB rats
PPI (top) and ASR (bottom) 7SEM in rats treated with
clozapine. Significantly different from vehicle-treated LE rats represented by**(Po0.01). Significantly different from vehicle-treated BB rats representedby # (Po0.05).
PPI (top) and ASR (bottom) 7SEM in rats treated with
PD149169. Significantly different from vehicle-treated LE rats representedby **(Po0.01). Significantly different from vehicle-treated BB rats
ence in the ASR between BB and LE rats, but PD149163
represented by # (Po0.05) and ## (Po0.01).
significantly decreased the ASR in both strains of rat asevidenced by a main effect of PD149163 (F(3,68) ¼ 5.3,Po0.01). There was no significant strain  PD149163
ASR in BB rats and this effect was also reported by another
interaction effect. Each dose of PD149163 significantly
group (Warren and Gash, 1983). The current data are only
reduced (Po0.05) the ASR in BB rats compared to vehicle,
partially consistent with those reports since we observed a
whereas the highest dose (3 mg/kg) reduced the ASR in LE
significantly higher startle response in BB rats in the
haloperidol experiment but not in the clozapine orPD149163 experiment. The reason for this variability inthe ASR strain differences is not clear; however, it indicates
that there is a disassociation between the ASR straindifferences and the PPI strain differences, an observation we
Figure 4 illustrates the catalepsy findings. There was a main
also made in our original report. This is also supported by
effect of drug treatment (F(5,42) ¼ 13.6, Po0.001). Post hoc
the fact that all three drugs tended to decrease ASR in LE
comparisons indicated that haloperidol (Po0.001) but no
and BB rats with efficacy that did not correspond to their
dose of clozapine or PD149163 significantly increased the
Acute administration of haloperidol did not affect PPI in
either LE or BB rats, which is also consistent with ourprevious report. Control (vehicle-treated) rats in the
haloperidol group exhibited PPI that was higher (approxi-
Consistent with our previous report (Feifel and Priebe,
mately 40%) than the control groups in the clozapine and
2001), untreated (vehicle) BB rats exhibited reduced PPI
PD149163 experiments (approximately 30%). However, it is
compared to untreated LE rats in each of the three
unlikely that this difference contributed to the negative drug
experiments. In our previous study, we reported higher
effects in the haloperidol experiment since an analysis of
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
phrenia (ie hallucinations, delusions), but they are lessefficacious against ‘negative’ symptoms (ie paucity ofthought, decreased emotional expression, decreased voli-
tion behavior) or the cognitive deficits known to be
associated with the disease (Meltzer, 2002). More recently,a second generation of antipsychotics has been developed.
These ‘atypical’ antipsychotics, of which clozapine is the
prototype, appears to be more efficacious, particularly in
ameliorating negative symptoms and cognitive deficitsassociated with schizophrenia (Kinon and Lieberman,
1996). Preclinical paradigms that can differentiate atypical
from typical clinical profiles are needed since advances in
the therapeutic field make it no longer desirable to develop
compounds with first generation clinical profiles. Since it is
established that atypical antipsychotics can produce desir-able clinical effects not associated with typical antipsycho-
tics, it is reasonable to assume that there exist preclinical
paradigms that model this atypical antipsychotic clinicaladvantage. Thus, whereas sensitivity to haloperidol wasonce considered the ‘litmus test’ for validating preclinicalmodels of antipsychotic potential, it is now desirable todevelop preclinical models that are preferentially sensitiveto atypical antipsychotics over typical antipsychotics suchas haloperidol. Indeed, several preclinical paradigms have
Catalepsy scored as the time paws of rats remained on the bar
been proposed to be useful for identifying putative
in seconds (7SEM). Significantly different from saline group represented by
antipsychotics of atypical category. Typically in these
paradigms, second-generation antipsychotics have a spec-trum of effects that is distinct or broader than typical
PPI produced by the weakest prepulse tested (4 dB), which
antipsychotics. Examples of these effects include antagon-
produced a mean PPI of 26% in the control group, was not
ism of PPI disruption produced by NMDA antagonists
increased by any of the haloperidol doses (PPI of 25%, 28%,
(Geyer et al, 2001) and induction of a distinct regional
and 28% for low, mid and high dose haloperidol,
pattern of immediate early gene expression (Deutch and
Duman, 1996). In this respect, it is significant that clozapine
In our earlier study, we tested only a single dose of
was able to reverse PPI deficits in BB rats. This reversal was
haloperidol (0.5 mg/kg) and therefore we could not rule out
dose dependent with the 10 and 15 mg/kg dose exhibiting
the possibility that higher or lower doses would be more
the greatest efficacy. It is not likely that clozapine’s ability to
effective. In this experiment, higher and lower doses in
acutely reverse PPI deficits in BB rats is due to a non-
addition to the 0.5 mg/kg dose were tested. Since none of the
specific pharmacological effect not associated with its
doses of haloperidol had a significant effect, we can
therapeutic mechanism, sedation for example. Whereas
reasonably conclude that the PPI deficit in BB rats is not
nonspecific effects such as sedation typically reduce normal
affected by acute administration of haloperidol. However, it
behavior, for example, locomotor activity, and can thus
is known from our previous study that chronic haloperidol
appear similar to the specific pharmacological effects of
reverses PPI deficits in BB rats (Feifel and Priebe, 2001). The
antipsychotics, restoration of deficient process, particularly
lack of an effect after acute administration of any dose of
an information processing deficit such as PPI, is unlikely to
haloperidol in this study confirms that differences in
be produced by a nonspecific effect. Consistent with this
efficacy between chronic and acute haloperidol treatment
notion, Depoortere et al (1997) concluded that clozapine’s
are due to the temporal aspects of the two treatments, rather
enhancement of PPI was not likely due to its sedating
properties since sedating psychotropic drugs that do not
Haloperidol is a prototype of the first generation, or
have antipsychotic properties, for example, diazepam,
‘typical’ family of antipsychotic drugs. These antipsychotics
decrease rather than facilitate PPI (Depoortere et al, 1997).
produce their therapeutic effects via a single pharmac-
The fact that clozapine but not haloperidol reversed PPI
ological mechanism, antagonism of D2 receptors (Feldman
deficits in BB rats suggests that acute reversal of PPI deficits
and Quenzer, 1997). Haloperidol’s lack of efficacy also
in BB rats may also have utility as a predictive model for
indicates that excessive dopamine transmission is not likely
atypical antipsychotic-like properties among putative anti-
to fully account for the PPI deficits seen in BB rats. Typical
psychotics. While there is some debate as to what
antipsychotics have notable limitations with respect to
pharmacological properties underlie the clinical advantages
clinical efficacy. For example, a significant proportion of
associated with atypical antipsychotics, there is general
schizophrenia patients fail to respond adequately to
agreement that combination of antagonism at both the
haloperidol and other antipsychotics from the typical family
5HT2A and D2 receptors is vital (Meltzer, 2002). The
(Stern et al, 1994). Furthermore, experience with haloper-
pattern of PPI effects observed with haloperidol and
idol and other typical antipsychotics indicates that they
clozapine in this and other studies may be understood if
have good efficacy against ‘positive’ symptoms of schizo-
inhibition of dopamine-2 transmission is sufficient to
Reversal of sensorimotor gating deficits in BB rats
reverse BB deficits chronically, and that other pharmaco-
effects of the NMDA antagonist, MK801 (Feifel et al, 1999),
logical mechanisms, for example, 5HT2 antagonism, po-
and DOI, a 5HT2A agonist (Feifel et al, 2003). These results
tentiate the D2 inhibition to produce a stronger, and thus
suggested that PD149163 produces antipsychotic-like pre-
more rapid reversal of PPI deficits. In this way, inhibition of
clinical effects by mechanisms other than inhibition of
D2 transmission may be necessary and sufficient for
chronic reversal of PPI deficits in BB rats and necessary
In the current study, PD149163 produced a very distinct
but not sufficient for acute reversal.
reversal of PPI deficits in BB rats. In fact, after treatment
However, other explanations are possible. For example, in
with the lowest dose, PPI in BB rats was higher than LE rats,
addition to sharing clinical properties with all other atypical
although the difference did not reach statistical significance.
antipsychotics, clozapine is also considered by many
The current results also provide further evidence that
clinicians and investigators to be singular among anti-
indirect inhibition of dopamine transmission cannot
psychotics in regards to efficacy (Taylor and Duncan-
account, by itself, for the antipsychotic-like effects of
McConnell, 2000; Chakos et al, 2001; Conley and Kelly,
PD149163, since PD149163 was effective in reversing BB
2001). This is particularly evident in the high success rates
rat PPI deficits, whereas haloperidol, a potent D2 antago-
with clozapine among treatment-resistant patients with
nist, was not. In this study, PD149163’s effect on BB PPI was
schizophrenia (Kane et al, 1988; Chakos et al, 2001; Kane
more consistent with clozapine, than with haloperidol.
et al, 2001). Thus, rather than due to mechanisms shared by
While this finding does not by itself suggest that PD149163
all atypical antipsychotics (eg D2 and 5HT2 antagonism),
has potential as an antipsychotic, the current finding is
clozapine’s ability to abolish PPI deficits in BB rats may be
consistent with previous evidence suggesting that PD149163
due to a putative mechanism which distinguishes clozapine
has a preclinical profile consistent with atypical anti-
from all other currently available antipsychotics, and which
psychotic drugs (Feifel et al, 2003). In this respect,
is responsible for its unique efficacy profile. If this is the
PD149163’s effects on PPI in BB rats supports the notion
case, acute reversal of PPI deficits in BB rats may be a model
that the BB rat is a predictive model for antipsychotics with
for drugs with novel properties that, like clozapine, are
atypical or novel mechanisms. The fact that PD149163, like
associated with a superior level of efficacy to current typical
clozapine, did not produce significant catalepsy, whereas
and atypical antipsychotics. Studies with other typical and
haloperidol did, supports the contention that PD149163 has
atypical antipsychotic drugs will be important in order to
a profile more similar to atypical than typical antipsychotics
determine whether PPI deficits in the BB rats is a predictive
and strengthens the evidence that acute PPI reversal in BB
model of the clinical effects associated with atypical
rats has predictive validity for drugs with atypical
antipsychotics or of novel drugs useful in treatment
In terms of establishing the validity of the BB model as
PD149163 appeared to produce the most robust effects on
predictive screen for atypical antipsychotic drugs, these
PPI reversing PPI strain differences at all doses tested even
findings should be considered preliminary. Additional
though it appeared to produce a nonsignificant tendency to
studies with other antipsychotics in the BB model will be
increase PPI in LE rats. PD149163’s ability to reverse the PPI
needed to address the issue of whether PD149163’s effects
deficits in BB rats is consistent with the notion that the BB
are shared by other atypical antipsychotics and are thus
rat is a useful predictive model of antipsychotic efficacy.
suggestive of ‘atypical’ clinical features or whether
Significant evidence exists that neurotensin, a neuropeptide,
PD149163’s effects are shared only by clozapine and are
may act as an endogenous antipsychotic and that it may, in
thus suggestive of a uniquely superior efficacy similar to
fact, mediate some of the clinical effects of antipsychotic
clozapine. It is noteworthy that PD149163 produced the
drugs (Kinkead and Nemeroff, 2002). There is also a large
most robust reversal of PPI deficits in BB rats of the three
body of evidence that neurotensin and neurotensin agonists
compounds tested, a finding that is auspicious for the
produce antipsychotic-like effects in preclinical studies.
therapeutic potential of drugs that target neurotensin
This has led many investigators to propose that neurotensin
agonists may have clinical potential as antipsychotic drugs
In summary, the BB rat offers a model of sensorimotor
(for a review, see Caceda et al, 2003). We have previously
gating deficits and a predictive model of antipsychotic
shown that administration of neurotensin produces anti-
potential with many novel and useful features. First, this
psychotic-like effects on PPI (Feifel et al, 1997). The
model does not require pharmacological, environmental, or
mechanism implicated in the antipsychotic-like effects
neuroanotomical manipulations to produce PPI deficits.
produced by neurotensin has been inhibition of dopamine
Rather, PPI deficits homologous to those seen in schizo-
transmission in the mesolimbic pathway, although neuro-
phrenia and other neuropsychiatric disorders are exhibited
tensin does not have significant affinity for dopamine
spontaneously and presumably due to the single gene
receptors (Adachi et al, 1990; Nouel et al, 1992). The 8–13
abnormality associated with these rats. As a genetic model
amino-acid fragment of neurotensin is the smallest
of PPI deficits, the BB rat exhibits greater construct validity
fragment that retains full biological activity of the parent
for the neuropsychiatric disorders associated with PPI
tridecapeptide (Kanba et al, 1988). PD149163 was developed
deficits compared to models requiring pharmacological,
by modifying neurotensin (8–13) to make it more stable to
environmental, or neuroanotomical manipulations to pro-
endopeptidase degradation and has been shown to cross the
duce PPI deficits. This improved construct validity affords
blood–brain barrier after parenteral administration (Wus-
the possibility of using the BB rats to explore the
trow et al, 1995). We have previously shown that PD149163
neurobiological and genetic substrates underlying sensori-
antagonizes amphetamine-induced disruption of PPI (Feifel
motor gating abnormalities that may also underlie the
et al, 1999). PD149163 also antagonized the PPI disrupting
neuropsychiatric conditions associated with such deficits.
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
As in the context of acute administration, the BB rat model
Feifel D, Minor KL, Dulawa S, Swerdlow NR (1997). The effects of
appears to differentiate typical from nontypical antipsycho-
intra-accumbens neurotensin on sensorimotor gating. Brain Res
tics and/or drugs with clozapine-like efficacy from drugs
with more conventional efficacy, it may also be a useful
Feifel D, Priebe K (2001). Vasopressin-deficient rats exhibit
predictive screen for novel antipsychotic drugs. Haloper-
sensorimotor gating deficits that are reversed by subchronic
idol’s effect on BB PPI is greater after chronic administra-
haloperidol. Biol Psychiatry 50: 425–433.
Feifel D, Reza TL, Wustrow DJ, Davis MD (1999). Novel
tion than after acute administration (Feifel and Priebe,
antipsychotic-like effects on prepulse inhibition of startle
2001). Therefore, the BB rat model may also be useful for
produced by a neurotensin agonist. J Pharmacol Exp Therapeut
elucidating the mechanisms underlying the therapeutic time
course, which is typically associated with antipsychotic
Feldman RS MJ, Quenzer LF (1997). Principles of Neuropsycho-
treatment in schizophrenia patients. In this regard, it will be
pharmacology. Sinauer Associates, Inc.: Sunderland, MA.
important to examine how the effects of chronic adminis-
Geyer MA, Krebs-Thomson K, Braff DL, Swerdlow NR (2001).
tration of clozapine and PD149163 compare to the effect of
Pharmacological studies of prepulse inhibition models of
acute administration of these drugs on BB PPI deficits.
sensorimotor gating deficits in schizophrenia: a decade inreview. Psychopharmacology (Berl) 156: 117–154.
Geyer MA, Wilkinson LS, Humby T, Robbins TW (1993). Isolation
rearing of rats produces a deficit in prepulse inhibition ofacoustic startle similar to that in schizophrenia. Biologic
DF is supported by a NIMH grant (MH62451). We
are grateful for the NIMH Chemical Synthesis Program
Kanba KS, Kanba S, Nelson A, Okazaki H, Richelson E (1988).
and SRI international for providing PD149163. We thank
[3H]neurotensin(8–13) binds in human brain to the same sites as
Dr. Neal Swerdlow for assistance with the catalepsy
does [3H]neurotensin but with higher affinity. J Neurochem 50:131–137.
Kane JM, Honigfeld G, Singer J, Meltzer H (1988). Clozapine in
treatment-resistant schizophrenics. Psychopharmacol Bull 24:
Kane JM, Marder SR, Schooler NR, Wirshing WC, Umbricht D,
Adachi DK, Kalivas PW, Schenk JO (1990). Neurotensin binding to
Baker RW et al (2001). Clozapine and haloperidol in moderately
dopamine. J Neurochem 54: 1321–1328.
refractory schizophrenia: a 6-month randomized and double-
Braff DL, Geyer MA (1990). Sensorimotor gating and schizo-
blind comparison. Arch Gen Psychiatry 58: 965–972.
phrenia. Human and animal model studies. Arch Gen Psychiatry
Kinkead B, Nemeroff CB (2002). Neurotensin: an endogenous
antipsychotic? Curr Opin Pharmacol 2: 99–103.
Caceda R, Kinkead B, Nemeroff CB (2003). Do neurotensin
Kinon BJ, Lieberman JA (1996). Mechanisms of action of atypical
receptor agonists represent a novel class of antipsychotic drugs?
antipsychotic drugs: a critical analysis. Psychopharmacology
Semin Clin Neuropsychiatry 8: 94–108.
Chakos M, Lieberman J, Hoffman E, Bradford D, Sheitman B
Laycock JF, Gartside IB, Chapman JT (1983). A comparison of the
(2001). Effectiveness of second-generation antipsychotics in
learning abilities of Brattleboro rats with hereditary diabetes
patients with treatment-resistant schizophrenia: a review and
insipidus and Long-Evans rats using positively reinforced
meta-analysis of randomized trials. Am J Psychiatry 158:
operant conditioning. Prog Brain Res 60: 183–187.
Le Pen G, Moreau JL (2002). Disruption of prepulse inhibition of
Cilia J, Reavill C, Hagan JJ, Jones DN (2001). Long-term evaluation
startle reflex in a neurodevelopmental model of schizophrenia:
of isolation-rearing induced prepulse inhibition deficits in rats.
reversal by clozapine, olanzapine and risperidone but not by
Psychopharmacology (Berl) 156: 327–337.
haloperidol. Neuropsychopharmacology 27: 1–11.
Conley RR, Kelly DL (2001). Management of treatment resistance
Lipska BK, Swerdlow NR, Geyer MA, Jaskiw GE, Braff DL,
in schizophrenia. Biol Psychiatry 50: 898–911.
Weinberger DR (1995). Neonatal excitotoxic hippocampal
Costall B, Hui SC, Naylor RJ (1978). Correlation between multitest
damage in rats causes post-pubertal changes in prepulse
and single test catalepsy assessment. Neuropharmacology 17:
inhibition of startle and its disruption by apomorphine.
Depoortere R, Perrault G, Sanger DJ (1997). Potentiation of
Mansbach RS, Geyer MA (1989). Effects of phencyclidine and
prepulse inhibition of the startle reflex in rats: pharmacological
phencyclidine biologs on sensorimotor gating in the rat.
evaluation of the procedure as a model for detecting anti-
Neuropsychopharmacology 2: 299–308.
psychotic activity. Psychopharmacology 132: 366–374.
Mansbach RS, Geyer MA, Braff DL (1988). Dopaminergic
Deutch AY, Duman RS (1996). The effects of antipsychotic drugs
stimulation disrupts sensorimotor gating in the rat. Psychophar-
on Fos protein expression in the prefrontal cortex: cellular
localization and pharmacological characterization. Neuroscience
Meltzer HY (2002). Mechanism of action of atypical antipsychotic
drugs. In: Davis KLC, Joseph Coyle D, Nemeroff C (eds).
Engelmann M, Landgraf R (1994). Microdialysis administration of
Neuropsychopharmacology: The Fifth Generation of Progress.
vasopressin into the septum improves social recognition in
American College of Neuropsychopharmacology, Lippincot,
Brattleboro rats. Physiol Behav 55: 145–149.
Feenstra MG, Snijdewint FG, Van Galen H, Boer GJ (1990).
Nouel D, Costentin J, Lugrin D, Kitabgi P, Ple N, Davoust D (1992).
Widespread alterations in central noradrenaline, dopamine, and
Investigations about a direct neurotensin-dopamine interaction
serotonin systems in the Brattleboro rat not related to the local
by nuclear magnetic resonance study, synaptosomal uptake of
absence of vasopressin. Neurochem Res 15: 283–288.
dopamine, and binding of neurotensin to its receptors.
Feifel D, Melendez G, Shilling PD (2003). A systemically
administered neurotensin agonist blocks disruption of prepulse
Sipes TA, Geyer MA (1994). Multiple serotonin receptor subtypes
inhibition produced by a serotonin-2A agonist. Neuropsycho-
modulate prepulse inhibition of the startle response in rats.
Reversal of sensorimotor gating deficits in BB rats
Stanley ME, Glick SD (1976). Interaction of drug effects with
Warren PH, Gash DM (1983). Hyperreflexive behavior in
testing procedures in the measurement of catalepsy. Neurophar-
Brattleboro rats. Peptides 4: 421–424.
Weiss IC, Feldon J, Domeney AM (1999). Circadian time does not
Stern RG, Kahn RS, Davidson M, Nora RM, Davis KL (1994). Early
modify the prepulse inhibition response or its attenuation by
response to clozapine in schizophrenia. Am J Psychiatry 151:
apomorphine. Pharmacol, Biochem Behav 64: 501–505.
Williams AR, Carey RJ, Miller M (1983). Behavioral differences
Swerdlow NR, Geyer MA (1998). Using an animal model of
between vasopressin-deficient (Brattleboro) and normal Long-
deficient sensorimotor gating to study the pathophysiology and
new treatments of schizophrenia. Schizophrenia Bulletin 24:
Williams AR, Carey RJ, Miller M (1985). Altered emotionality of
the vasopressin-deficient Brattleboro rat. Peptides 6(Suppl 1):
Taylor DM, Duncan-McConnell D (2000). Refractory schizo-
phrenia and atypical antipsychotics. J Psychopharmacol 14:
Wustrow DJ DM, Akunne HC, Corbin AE, Wiley JN, Wise LD,
Heffner TG (1995). Reduced amide bond neurotensin 8-13
Wadenberg ML (1996). Serotonergic mechanisms in neuroleptic-
mimetics with potent in vivo activity. Bioorgan Medicin Chem
induced catalepsy in the rat. Neurosci Biobehav Rev 20: 325–339.
PERTUSSIS (WHOOPING COUGH) EXPOSURE NOTICE One or more children at your child’s school have been diagnosed with pertussis (whooping cough). These students are currently being treated with antibiotics and will remain at home until no longer infectious, however, your child may have been exposed to pertussis while at school. We are sending you this letter to make you aware of what symptoms
(A more detailed list of contents is on the campus). Unit 1: Characteristics and classification of living organisms - List and desrcibe the 7 characteristics of living organisms. - Define the bionomial system of classification - Recognize the main features of: bony fish, amphibians, reptiles, birds, mammals, Flowering plants (monocotyledons vs dicotyledons), crustaceans, arachnids, myriapod
 Neuropsychopharmacology (2004) 29, 731–738& 2004 Nature Publishing Group
Reversal of Sensorimotor Gating Deficits in Brattleboro Ratsby Acute Administration of Clozapine and a NeurotensinAgonist, but not Haloperidol: a Potential Predictive Model forNovel Antipsychotic Effects
David Feifel*,1, Gilia Melendez1 and Paul D Shilling1
Department of Psychiatry, University of California, San Diego, San Diego, CA, USA
Prepulse inhibition (PPI) of acoustic startle is decreased in unmedicated schizophrenia patients and similar deficits can be induced in rats
through pharmacological, environmental, or neuroanatomical manipulations. Recently, we reported that Brattleboro (BB) rats, a Long
Evans (LE) strain with a single gene mutation, have inherent deficits in PPI homologous to those observed in schizophrenia patients. We
also reported that PPI deficits in BB rats could be reversed by chronic but not acute administration of 0.5 mg/kg haloperidol. No other
dose or drug was tested in that experiment. In this study, we tested the effects of acute subcutaneous administration of several doses of
haloperidol as well as the second-generation antipsychotic, clozapine, and the putative novel antipsychotic, PD149163, a neurotensin
mimetic that crosses the blood–brain barrier. Consistent with our previous report, BB rats exhibited PPI deficits compared to LE rats and
none of the doses of haloperidol produced a significant effect on this PPI deficit. In contrast, 10 and 15 mg/kg of clozapine and all the
doses of PD149163 tested reversed the PPI deficits in BB rats. In addition, haloperidol, but not clozapine or PD149163 produced
significant catalepsy in BB rats, supporting the notion that PD149163 has a profile consistent with atypical antipsychotics and providing
support for the predictive validity of the PPI results. These results further strengthen the notion that the BB rat is a useful predictive
model of antipsychotic efficacy and suggest that this model may differentiate between antipsychotics belonging to different therapeutic
categories, for example, first- and second-generation agents.
Neuropsychopharmacology (2004) 29, 731–738& 2004 Nature Publishing Group
Reversal of Sensorimotor Gating Deficits in Brattleboro Ratsby Acute Administration of Clozapine and a NeurotensinAgonist, but not Haloperidol: a Potential Predictive Model forNovel Antipsychotic Effects
David Feifel*,1, Gilia Melendez1 and Paul D Shilling1
Department of Psychiatry, University of California, San Diego, San Diego, CA, USA
Prepulse inhibition (PPI) of acoustic startle is decreased in unmedicated schizophrenia patients and similar deficits can be induced in rats
through pharmacological, environmental, or neuroanatomical manipulations. Recently, we reported that Brattleboro (BB) rats, a Long
Evans (LE) strain with a single gene mutation, have inherent deficits in PPI homologous to those observed in schizophrenia patients. We
also reported that PPI deficits in BB rats could be reversed by chronic but not acute administration of 0.5 mg/kg haloperidol. No other
dose or drug was tested in that experiment. In this study, we tested the effects of acute subcutaneous administration of several doses of
haloperidol as well as the second-generation antipsychotic, clozapine, and the putative novel antipsychotic, PD149163, a neurotensin
mimetic that crosses the blood–brain barrier. Consistent with our previous report, BB rats exhibited PPI deficits compared to LE rats and
none of the doses of haloperidol produced a significant effect on this PPI deficit. In contrast, 10 and 15 mg/kg of clozapine and all the
doses of PD149163 tested reversed the PPI deficits in BB rats. In addition, haloperidol, but not clozapine or PD149163 produced
significant catalepsy in BB rats, supporting the notion that PD149163 has a profile consistent with atypical antipsychotics and providing
support for the predictive validity of the PPI results. These results further strengthen the notion that the BB rat is a useful predictive
model of antipsychotic efficacy and suggest that this model may differentiate between antipsychotics belonging to different therapeutic
categories, for example, first- and second-generation agents. Reversal of sensorimotor gating deficits in BB rats
antipsychotic potential. Therefore, there is significant
effects on PPI are stable across the circadian cycle (Weiss
interest in development of nonpharmacological paradigms
et al, 1999). The rats were tested in startle chambers to
of PPI deficits. Two of the most studied examples of
characterize their baseline PPI and startle. Animals
nonpharmacological paradigms of PPI deficits are social
were assigned, based on their baseline PPI, to one of four
isolation-rearing (Geyer et al, 1993) and neonatal hippo-
groups matched so as to achieve comparable average PPI
campal lesions (Lipska et al, 1995). PPI deficits produced by
across groups. Drug treatment began 3 days after baseline
social isolation rearing are reversed by both typical and
atypical antipsychotics (Cilia et al, 2001; Le Pen and
In one experiment 50 LE rats and 50 BB rats were
Moreau, 2002). Preliminary findings suggest that PPI
administered subcutaneous (s.c.) injections of either 0
deficits produced by neonatal hippocampal lesions may be
(vehicle), 0.1, 0.5, or 1 mg/kg of haloperidol (UCSD Medical
reversed by atypical but not typical antipsychotics (Le Pen
Center, San Diego, CA). In another experiment 51 LE and 49
BB rats were administered s.c. injection of either 0 (vehicle),
Brattleboro (BB) rats are Long Evans (LE) rats with a
5, 10, or 15 mg/kg clozapine (Sigma Chemicals, St Louis,
single base pair mutation that results in the inability to
MO). In a third study, 43 LE rats and 33 BB rats were
properly synthesize the neurotransmitter and neurohor-
administered s.c. injection of either 0 (vehicle), 0.5, 1, or
mone vasopressin. BB rats have many behavioral and
2 mg/kg PD149163 (LYS(CH2NH)LYS-PRO-TRP-tLE-LEU-
cognitive abnormalities, including deficits in memory
OEt) (SRI International, NIMH chemical synthesis pro-
(Laycock et al, 1983), emotion (Williams et al, 1985), social
gram). Doses were selected based on their demonstrated
recognition (Engelmann and Landgraf, 1994), motivation,
ability to reverse PPI deficits in other reports (Feifel et al,
and attention (Williams et al, 1983). They have abnormal-
1999; Geyer et al, 2001). Vehicle for haloperidol was distilled
ities in brain systems including dopamine and serotonin,
water and the volume injected was 1 ml/kg. Vehicle for
neurotransmitters implicated in schizophrenia (Feenstra
clozapine and PD149163 was 0.1 N HCl and half volume
et al, 1990). BB rats also have deficits in PPI compared with
0.9% saline brought to pH 5–6 with a few drops of 1 N
their wild-type, LE counterparts. We previously showed that
NaOH. The volume injected for clozapine was 1.5 ml/kg and
acute treatment with a single dose (0.5 mg/kg) of haloper-
for PD149163 was 1 ml/kg. Each treatment group had a
idol did not reverse the PPI deficits in BB rats. In contrast,
chronic treatment with that dose of haloperidol reverses the
Animals were tested in startle chambers (San Diego
deficits, suggesting that the BB rat may be a very useful
Instruments, San Diego, CA) 20 min after drug administra-
genetic model of sensorimotor gating deficits associated
tion. Once placed in startle chambers, each rat had a 5-min
with schizophrenia that models the therapeutic time course
acclimation period. A 65-dB background noise was
of antipsychotic drugs (Feifel and Priebe, 2001). Since only
continuously present throughout the session. The acclima-
one dose of haloperidol was used in that study, it is possible
tion was followed by a 15 min PPI test session during which
that acute administration of higher or lower doses of
rats were presented with 40 ms 120 dB startle pulses without
haloperidol may reverse the PPI deficits observed in BB rats.
Reversal of sensorimotor gating deficits in BB rats
antipsychotic potential. Therefore, there is significant
effects on PPI are stable across the circadian cycle (Weiss
interest in development of nonpharmacological paradigms
et al, 1999). The rats were tested in startle chambers to
of PPI deficits. Two of the most studied examples of
characterize their baseline PPI and startle. Animals
nonpharmacological paradigms of PPI deficits are social
were assigned, based on their baseline PPI, to one of four
isolation-rearing (Geyer et al, 1993) and neonatal hippo-
groups matched so as to achieve comparable average PPI
campal lesions (Lipska et al, 1995). PPI deficits produced by
across groups. Drug treatment began 3 days after baseline
social isolation rearing are reversed by both typical and
atypical antipsychotics (Cilia et al, 2001; Le Pen and
In one experiment 50 LE rats and 50 BB rats were
Moreau, 2002). Preliminary findings suggest that PPI
administered subcutaneous (s.c.) injections of either 0
deficits produced by neonatal hippocampal lesions may be
(vehicle), 0.1, 0.5, or 1 mg/kg of haloperidol (UCSD Medical
reversed by atypical but not typical antipsychotics (Le Pen
Center, San Diego, CA). In another experiment 51 LE and 49
BB rats were administered s.c. injection of either 0 (vehicle),
Brattleboro (BB) rats are Long Evans (LE) rats with a
5, 10, or 15 mg/kg clozapine (Sigma Chemicals, St Louis,
single base pair mutation that results in the inability to
MO). In a third study, 43 LE rats and 33 BB rats were
properly synthesize the neurotransmitter and neurohor-
administered s.c. injection of either 0 (vehicle), 0.5, 1, or
mone vasopressin. BB rats have many behavioral and
2 mg/kg PD149163 (LYS(CH2NH)LYS-PRO-TRP-tLE-LEU-
cognitive abnormalities, including deficits in memory
OEt) (SRI International, NIMH chemical synthesis pro-
(Laycock et al, 1983), emotion (Williams et al, 1985), social
gram). Doses were selected based on their demonstrated
recognition (Engelmann and Landgraf, 1994), motivation,
ability to reverse PPI deficits in other reports (Feifel et al,
and attention (Williams et al, 1983). They have abnormal-
1999; Geyer et al, 2001). Vehicle for haloperidol was distilled
ities in brain systems including dopamine and serotonin,
water and the volume injected was 1 ml/kg. Vehicle for
neurotransmitters implicated in schizophrenia (Feenstra
clozapine and PD149163 was 0.1 N HCl and half volume
et al, 1990). BB rats also have deficits in PPI compared with
0.9% saline brought to pH 5–6 with a few drops of 1 N
their wild-type, LE counterparts. We previously showed that
NaOH. The volume injected for clozapine was 1.5 ml/kg and
acute treatment with a single dose (0.5 mg/kg) of haloper-
for PD149163 was 1 ml/kg. Each treatment group had a
idol did not reverse the PPI deficits in BB rats. In contrast,
chronic treatment with that dose of haloperidol reverses the
Animals were tested in startle chambers (San Diego
deficits, suggesting that the BB rat may be a very useful
Instruments, San Diego, CA) 20 min after drug administra-
genetic model of sensorimotor gating deficits associated
tion. Once placed in startle chambers, each rat had a 5-min
with schizophrenia that models the therapeutic time course
acclimation period. A 65-dB background noise was
of antipsychotic drugs (Feifel and Priebe, 2001). Since only
continuously present throughout the session. The acclima-
one dose of haloperidol was used in that study, it is possible
tion was followed by a 15 min PPI test session during which
that acute administration of higher or lower doses of
rats were presented with 40 ms 120 dB startle pulses without
haloperidol may reverse the PPI deficits observed in BB rats. Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
1. PPI in untreated BB rats is significantly lower than in
2. Treatment with the test drugs facilitates PPI in BB rats
3. Treatment of BB rats with the test drugs restores their
Data of the acoustic startle response (ASR) to the startle
stimuli not preceded by any prepulse were subjected to
analysis using a similar two factor ANOVA.
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
1. PPI in untreated BB rats is significantly lower than in
2. Treatment with the test drugs facilitates PPI in BB rats
3. Treatment of BB rats with the test drugs restores their
Data of the acoustic startle response (ASR) to the startle
stimuli not preceded by any prepulse were subjected to
analysis using a similar two factor ANOVA. Reversal of sensorimotor gating deficits in BB rats
PPI (top) and ASR (bottom) 7SEM in rats treated with
clozapine. Significantly different from vehicle-treated LE rats represented by**(Po0.01). Significantly different from vehicle-treated BB rats representedby # (Po0.05).
Reversal of sensorimotor gating deficits in BB rats
PPI (top) and ASR (bottom) 7SEM in rats treated with
clozapine. Significantly different from vehicle-treated LE rats represented by**(Po0.01). Significantly different from vehicle-treated BB rats representedby # (Po0.05).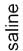

 Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
phrenia (ie hallucinations, delusions), but they are lessefficacious against ‘negative’ symptoms (ie paucity ofthought, decreased emotional expression, decreased voli-
tion behavior) or the cognitive deficits known to be
associated with the disease (Meltzer, 2002). More recently,a second generation of antipsychotics has been developed.
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
phrenia (ie hallucinations, delusions), but they are lessefficacious against ‘negative’ symptoms (ie paucity ofthought, decreased emotional expression, decreased voli-
tion behavior) or the cognitive deficits known to be
associated with the disease (Meltzer, 2002). More recently,a second generation of antipsychotics has been developed. Reversal of sensorimotor gating deficits in BB rats
reverse BB deficits chronically, and that other pharmaco-
effects of the NMDA antagonist, MK801 (Feifel et al, 1999),
logical mechanisms, for example, 5HT2 antagonism, po-
and DOI, a 5HT2A agonist (Feifel et al, 2003). These results
tentiate the D2 inhibition to produce a stronger, and thus
suggested that PD149163 produces antipsychotic-like pre-
more rapid reversal of PPI deficits. In this way, inhibition of
clinical effects by mechanisms other than inhibition of
D2 transmission may be necessary and sufficient for
chronic reversal of PPI deficits in BB rats and necessary
In the current study, PD149163 produced a very distinct
but not sufficient for acute reversal.
Reversal of sensorimotor gating deficits in BB rats
reverse BB deficits chronically, and that other pharmaco-
effects of the NMDA antagonist, MK801 (Feifel et al, 1999),
logical mechanisms, for example, 5HT2 antagonism, po-
and DOI, a 5HT2A agonist (Feifel et al, 2003). These results
tentiate the D2 inhibition to produce a stronger, and thus
suggested that PD149163 produces antipsychotic-like pre-
more rapid reversal of PPI deficits. In this way, inhibition of
clinical effects by mechanisms other than inhibition of
D2 transmission may be necessary and sufficient for
chronic reversal of PPI deficits in BB rats and necessary
In the current study, PD149163 produced a very distinct
but not sufficient for acute reversal. Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
As in the context of acute administration, the BB rat model
Feifel D, Minor KL, Dulawa S, Swerdlow NR (1997). The effects of
appears to differentiate typical from nontypical antipsycho-
intra-accumbens neurotensin on sensorimotor gating. Brain Res
tics and/or drugs with clozapine-like efficacy from drugs
with more conventional efficacy, it may also be a useful
Feifel D, Priebe K (2001). Vasopressin-deficient rats exhibit
predictive screen for novel antipsychotic drugs. Haloper-
sensorimotor gating deficits that are reversed by subchronic
idol’s effect on BB PPI is greater after chronic administra-
haloperidol. Biol Psychiatry 50: 425–433.
Reversal of sensorimotor gating deficits in BB ratsD Feifel et al
As in the context of acute administration, the BB rat model
Feifel D, Minor KL, Dulawa S, Swerdlow NR (1997). The effects of
appears to differentiate typical from nontypical antipsycho-
intra-accumbens neurotensin on sensorimotor gating. Brain Res
tics and/or drugs with clozapine-like efficacy from drugs
with more conventional efficacy, it may also be a useful
Feifel D, Priebe K (2001). Vasopressin-deficient rats exhibit
predictive screen for novel antipsychotic drugs. Haloper-
sensorimotor gating deficits that are reversed by subchronic
idol’s effect on BB PPI is greater after chronic administra-
haloperidol. Biol Psychiatry 50: 425–433. Reversal of sensorimotor gating deficits in BB rats
Stanley ME, Glick SD (1976). Interaction of drug effects with
Warren PH, Gash DM (1983). Hyperreflexive behavior in
testing procedures in the measurement of catalepsy. Neurophar-
Brattleboro rats. Peptides 4: 421–424.
Reversal of sensorimotor gating deficits in BB rats
Stanley ME, Glick SD (1976). Interaction of drug effects with
Warren PH, Gash DM (1983). Hyperreflexive behavior in
testing procedures in the measurement of catalepsy. Neurophar-
Brattleboro rats. Peptides 4: 421–424.