Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
No job name
Biochemistry 2001, 40, 4323-4331
P-Glycoprotein-Mediated Colchicine Resistance in Different Cell Lines Correlates
with the Effects of Colchicine on P-Glycoprotein Conformation†
Todd E. Druley,‡ Wilfred D. Stein,§ Adam Ruth,‡ and Igor B. Roninson*,‡
Department of Molecular Genetics, UniVersity of Illinois at Chicago, Chicago, Illinois 60607, andDepartment of Biological Chemistry, Hebrew UniVersity, Jerusalem, Israel 91904ReceiVed June 15, 2000; ReVised Manuscript ReceiVed NoVember 7, 2000
ABSTRACT: The multidrug transporter P-glycoprotein (Pgp) is an ATPase efflux pump for multiple cytotoxicagents, including vinblastine and colchicine. We have found that resistance to vinblastine but not tocolchicine in cell lines derived from different types of tissues and expressing the wild-type human Pgpcorrelates with the Pgp density. Vinblastine induces a conformational change in Pgp, evidenced by increasedreactivity with a conformation-sensitive monoclonal antibody UIC2, in all the tested cell lines. In contrast,colchicine increases the UIC2 reactivity in only some of the cell lines. In those lines where colchicinealone did not affect UIC2 reactivity, this drug was, however, able to reverse the vinblastine-induced increasein UIC2 reactivity. The magnitude of the increase in UIC2 reactivity in the presence of saturatingconcentrations of colchicine correlates with the relative ability of Pgp to confer colchicine resistance indifferent cell lines, suggesting the existence of some cell-specific factors that have a coordinate effect onthe ability of colchicine to induce conformational transitions and to be transported by Pgp. Colchicine,like vinblastine, reverses the decrease in UIC2 reactivity produced by nonhydrolyzable nucleotides, butunlike vinblastine, it does not reverse the effect of ATP at a high concentration. Colchicine, however,decreases the Hill number for the effect of ATP on the UIC2 reactivity from 2 to 1. Colchicine increasesthe UIC2 reactivity and reverses the effect of ATP in ATPase-deficient Pgp mutants, but not in the wild-type Pgp expressed in the same cellular background, suggesting that ATP hydrolysis counteracts the effectsof colchicine on the Pgp conformation.
The multidrug transporter Pgp1 is an ATPase efflux pump
that affect drug transport by Pgp have been identified (8), it
for multiple cytotoxic agents, responsible for the best-known
is not known whether external factors may also affect the
form of multidrug resistance in tumor cells (1, 2). Pgp is a
relative efficacy of drug transport by Pgp. Romsicki and
170 kDa glycoprotein consisting of two homologous halves,
Sharom (9) have shown that the relative binding affinity of
each containing a nucleotide-binding domain with NBS-
different drugs for purified Pgp in lipid mixtures was affected
carrying consensus Walker A and Walker B sequence motifs,
by the lipid composition, suggesting that Pgp-drug interac-
characteristic of the ATP-binding cassette (ABC) family of
tions may also vary among cell types with different
transport proteins (3), and a hydrophobic domain with six
transmembrane segments. It has been demonstrated throughchemical means (4) or by amino acid substitutions located
We have previously shown that the reactivity of Pgp
in either the N-terminal or C-terminal Walker A motif (5,
encoded by the human MDR1 gene with a conformation-
6) that both NBS must be intact for Pgp to hydrolyze ATP.
specific monoclonal antibody UIC2 is affected by different
The presence of Pgp transport substrates has been shown to
Pgp ligands (10, 11). Specifically, the UIC2 reactivity is
increase the rate of ATP hydrolysis by Pgp (5, 7). A mutation
decreased by different Pgp-binding nucleotides, whereas Pgp
that alters the relative ability of Pgp to confer resistance to
substrates, such as vinblastine, reverse this effect of nucle-
different drugs was also found to change the ability of such
otides and increase the UIC2 reactivity. While most of the
drugs to stimulate ATP hydrolysis (5). While many mutations
Pgp-transported drugs were found to increase the UIC2reactivity in intact Pgp-expressing cells, three of these
† This work was supported by NIH Grant R37CA40333 (I.B.R.).
substrates (colchicine, etoposide, and puromycin) failed to
W.D.S. was the recipient of a Yamagiwa-Yoshida Memorial Interna-
increase the UIC2 reactivity in the original survey (10). In
tional Cancer Study grant administered by the International Union
the study presented here, we have found that colchicine can
* To whom correspondence should be addressed: Department of
increase the UIC2 reactivity in some but not all Pgp-
Molecular Genetics (M/C 669), University of Illinois at Chicago, 900
expressing cell lines, and that the ability of colchicine to
South Ashland Ave., Chicago, IL 60607-7170. E-mail: roninson@uic.edu.
increase the UIC2 reactivity correlates with its relative ability
to be transported by Pgp in different cells. We have also
1 Abbreviations: Pgp, P-glycoprotein; FACS, fluorescence-activated
used the UIC2 reactivity shift assay to characterize the effects
cell sorter; ABC, ATP-binding cassette; NBS, nucleotide-binding site-
of colchicine in the presence of vinblastine, ATP, or
(s); AMP-PNP, 5′-adenylylimidodiphosphate; MIANS, 2-(4-maleimi-doanilino)naphthalene-6-sulfonic acid.
nonhydrolyzable nucleotides. The results of these assays
4324 Biochemistry, Vol. 40, No. 14, 2001
indicate that ATP hydrolysis by Pgp counteracts the effects
sites on 1 × 106 cells. Following addition of the primary
of colchicine on the Pgp conformation.
antibody, the reaction mixture was held at 37 °C for anadditional 30 min. Primary antibody reactions were stopped
EXPERIMENTAL PROCEDURES
by the addition of 5 mL of ice-cold PBS and the cells thencentrifuged for 5 min at 4 °C and 1500 rpm. Pellets were
Materials. ATP, AMP-PNP, vinblastine, colchicine, and
resuspended in 100 µL of PBS and 1% BSA buffer
propidium iodide were from Sigma. Staphylococcus aureus
containing 25 µg/mL goat anti-mouse FITC-conjugated
-toxin was purchased from List Biological Labs. The goat
secondary antibody. The reaction mixtures were left on ice,
anti-mouse IgG2a fluorescein isothiocyanate (FITC)-conju-
to prevent the function of Pgp, for 30 min before the reactions
gated secondary antibody was obtained from Caltag Labo-
were stopped by the addition of 5 mL of ice-cold PBS buffer
ratories. MRK16 monoclonal antibody was generously
and the mixtures centrifuged for 5 min at 4 °C and 1500
provided by T. Tsuruo (University of Tokyo, Tokyo, Japan).
rpm. Immediately prior to FACS analysis, each pellet was
The isolation and preparation of the monoclonal antibody
resuspended in 350-500 µL of ice-cold PBS and 1% BSA
UIC2 have been previously described (12).
buffer containing 1 µg/mL propidium iodide (PI) and left
Cell Lines. LMtk- murine fibroblast cell lines, KK-H, KK-
on ice. Two-color cytofluorometric analysis was performed
L, KM-H, MK-H, and MM, were derived after transfection
by acquiring at least 10 000 individual events using a Becton
with either the wild-type (KK) or mutant (KM, MK, and
Dickinson FacSort flow cytometer. Flow cytometric data
MM) forms of human MDR1 cDNA, followed by vinblastine
were analyzed by using the Becton Dickinson Information
selection or (in the case of MM) by FACS sorting for the
expression of human Pgp (ref 10 and unpublished data). TheMK, KM, and MM mutants contain amino acid substitutions
Data Analysis. We determined the kinetic parameters from
at either one (KM or MK) or both (MM) conserved lysine
the data using the SigmaPlot program. This gave us (1) the
residues in the Walker A motifs of the N-terminal or
maximum or minimum fluorescence (Fmax or Fmin, respec-
C-terminal NBS, K433M and K1076M, respectively.
tively) at asymptotically high or low levels of vinblastine,nucleotide, or vanadate, (2) the concentration of vinblastine,
HT1080-MDR1 cells constitute a population of HT1080
nucleotide, or vanadate at which half of the maximal change
human fibrosarcoma cells infected with a recombinant
retroviral vector carrying human MDR1 cDNA, with Pgp-
m), and (3) the Hill number, n. The
best-fit regression through the respective data points was
positive cells isolated with the FACS (13). 3T3-MDR1 cells
determined using the appropriate binding isotherm, as
were derived without cytotoxic selection from mouse NIH
follows. We used eq 1 below for cases where the fluores-
3T3 fibroblasts as described in ref 14. The KB-GRC1 cell
cence signal, F, increases with B, the concentration of ligand,
line was derived following transfection of KB-3-1 cells with
and eq 2 for those cases in which the fluorescence decreases,
human MDR1 cDNA and a single step of colchicine selection
The K562/i-S9 cell line was derived from human K562
leukemia cells by infection with a recombinant retrovirus
carrying the human MDR1 cDNA followed by subcloning
- F )K n]/(K n + Bn)
(without cytotoxic selection) and immunostaining for Pgp
(16). The multidrug-resistant CEM/VLB-100 cell line wasderived from human T-lymphoblastoid CEM leukemia cells
S. aureus R-Toxin Permeabilization. The concentration and
by multistep selection with vinblastine (17).
time course of S. aureus R-toxin necessary to yield an
All attached cell lines were maintained in 15 cm tissue
approximate 50% distribution between PI-positive and PI-
culture plates in Dulbecco’s modified Eagle’s medium
negative staining cells was determined for each cell line in
containing 10% fetal bovine serum, 1% glutamine, and a
preliminary experiments. An R-toxin concentration was
1% penicillin/streptomycin mixture. Leukemic cells that grow
chosen that was effective in 15-30 min at 37 °C. The
in suspension were maintained in 75 cm2 flasks in RPMI
approximate percentage of permeabilization was checked
1640 medium containing 10% fetal bovine serum, 1%
periodically under a light microscope via trypan blue staining
glutamate, and a 1% penicillin/streptomycin mixture.
and counting the percentage of blue cells per high power
Fluorescence-ActiVated Cell Sorting Assays. All primary
field. Permeabilization was performed in PBS and 1% BSA
antibody staining reactions were carried out in a final volume
buffer at a final volume of 100 µL containing 1 × 106 cells
of 200 µL containing 1 × 106 cells/reaction mixture in
at 37 °C. The reaction was stopped by the addition of 30
phosphate-buffered saline (PBS) with 1% bovine serum
mL of 37 °C PBS, and the cells were centrifuged for 5 min
albumin (BSA). Cells were reacted with their respective
at 1500 rpm and room temperature. Pellets were resuspended
nucleotides and/or drugs for 10-15 min at 37 °C prior to
in PBS containing 10 mM MgCl2 with or without the
the addition of the primary antibody. UIC2 was aliquoted,
indicated nucleotide at 37 °C for 15 min, and then each
heated at 48 °C for 24 h in a thermal cycler, and stored at 4
reaction proceeded through the FACS assay procedure as
°C prior to use. This treatment did not affect the reactivity
of the antibody to Pgp in the presence of substrate, but
Cytotoxicity Assays. For assays involving adherent cell
decreased the reactivity of the antibody to Pgp in the absence
lines, 250 cells were plated in triplicate in a 3 cm plate
of substrate, thus increasing the sensitivity of the assay (our
(Falcon) in 2-3 mL of drug-free Dulbecco’s modified
unpublished data). The primary antibody concentration that
Eagle’s medium containing 10% fetal bovine serum, 1%
was used (15 µg/mL) had been previously determined to be
glutamine, and a 1% penicillin/streptomycin mixture. After
a saturating antibody concentration for all available binding
24 h, the drug-free medium was aspirated and replaced with
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4325
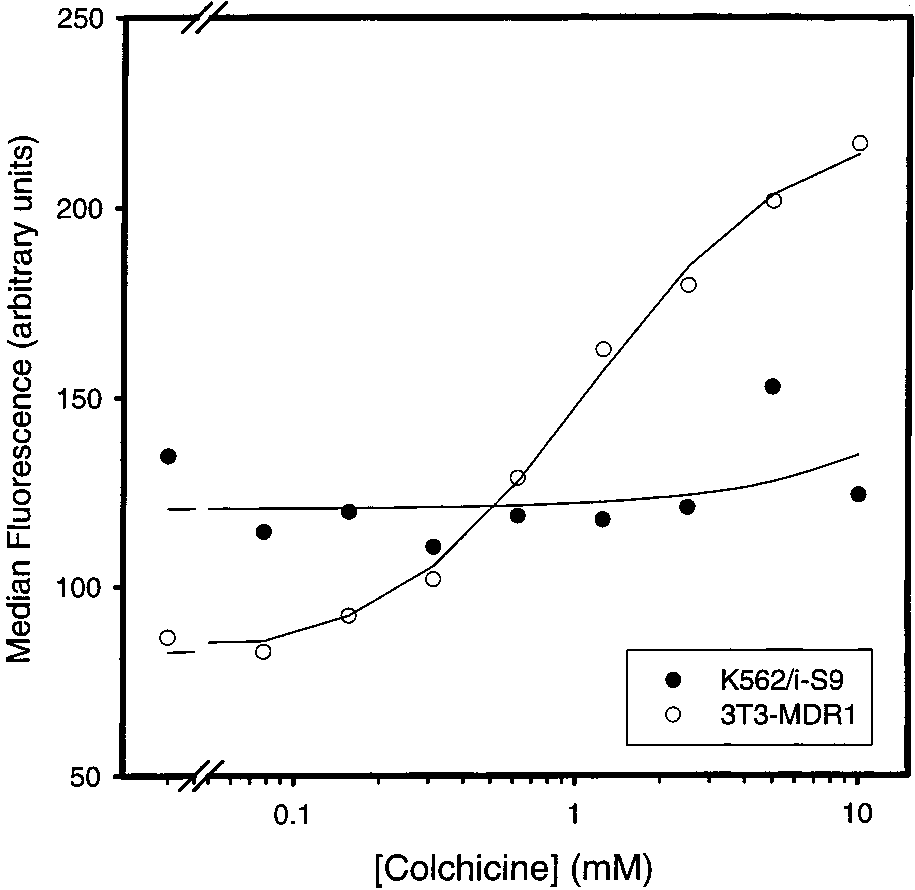
FIGURE 1: Effects of different concentrations of colchicine on UIC2reactivity in K562/i-S9 and 3T3-MDR1 cells. Intact K562/i-S9 (b)and 3T3-MDR1 (O) cells were incubated at the indicated concen-trations of colchicine for 10 min at 37 °C prior to the addition ofUIC2 for an additional 30 min at 37 °C.
fresh medium containing the appropriate type and concentra-tion of cytotoxic drug, in a 3 mL volume. After 8 days at 37
°C, the cells were fixed with methanol and stained withcrystal violet (10% w/v in a 10% methanol solution) and
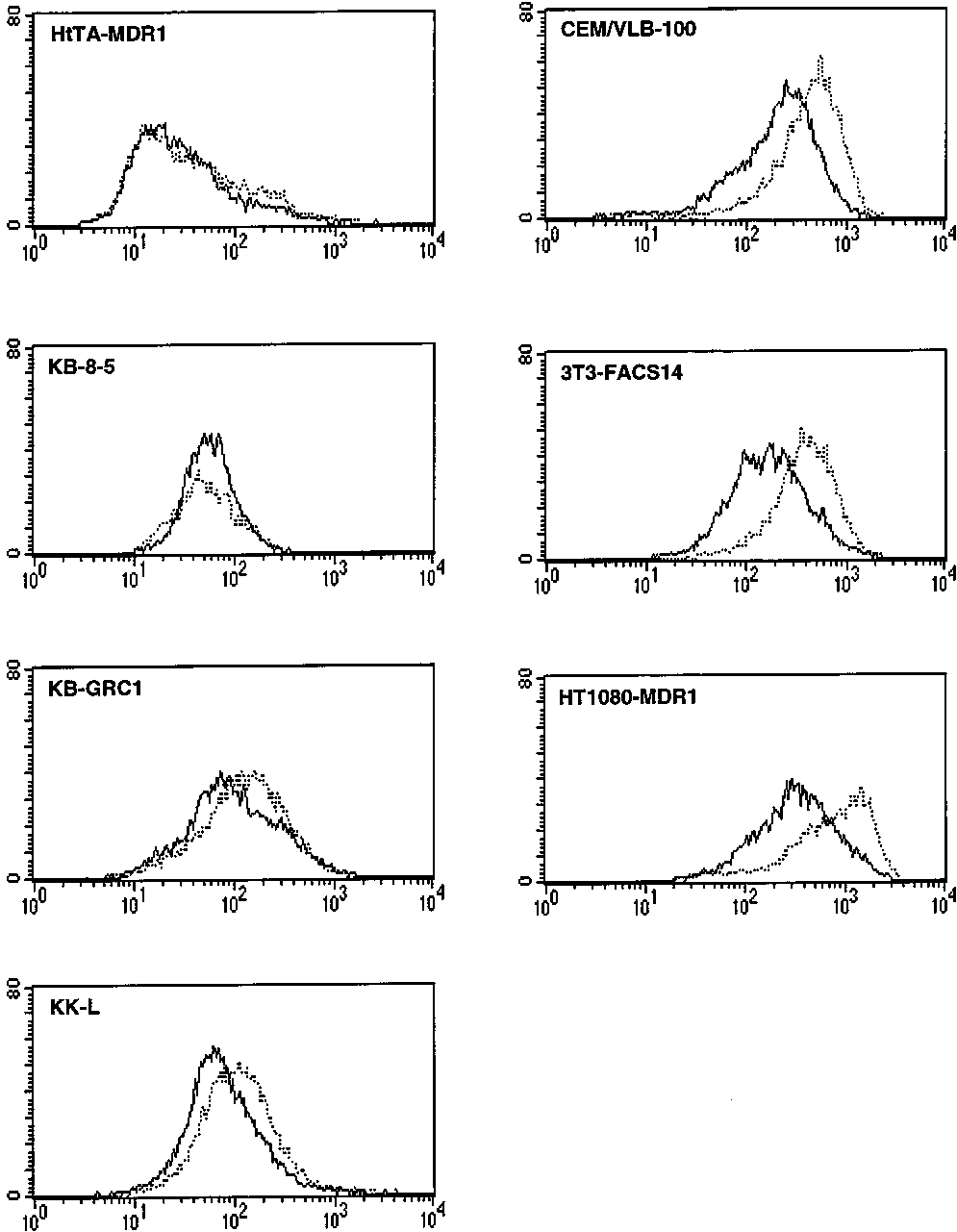
FIGURE 2: Effects of colchicine on UIC2 reactivity in different
the number of colonies per plate was counted.
cell types. Each histogram depicts the fluorescence intensity of theindicated cell type, all expressing wild-type human Pgp, stained
For assays with suspension cell lines, 3 mL of medium
with UIC2 in the absence (s) or presence (‚‚‚) of a saturating
containing 50 000 cells/mL and the appropriate concentration
concentration (5-10 mM) of colchicine.
of cytotoxic drug were plated in a 3 cm plate, in triplicate.
reactivity up to a concentration of 4% and increased this
The cells were left to incubate at 37 °C for 5-7 days. After
this incubation, cell clumps were disrupted by repeated
therefore account for the colchicine-induced increase in UIC2
pipetting and suspended in Isoton II Electrolyte Solution
reactivity in our reaction mixtures, which did not contain
(Coulter), and the cell number was determined using a
The solid line through the white circles in Figure 1 is the
best-fit regression as determined using eq 1 in ExperimentalProcedures. From this regression, the Km value for the effect
Differential Effects of Colchicine on the UIC2 ReactiVity
of colchicine on UIC2 reactivity in 3T3-MDR1 cells was
of the Human MDR1 Pgp in Different Cell Lines. Figure 1
1.11 ( 0.14 mM. The Km values determined via UIC2
depicts the effects of colchicine on the UIC2 reactivity of
reactivity assays are apparent affinities of the ligand for Pgp
Pgp in two multidrug-resistant cell lines. Due to the large
and not the intrinsic affinity of the ligand for the protein.
number of reaction conditions and controls necessary to
The Hill number (n) for the colchicine-induced increase in
determine accurate parameters of substrate interactions with
the UIC2 reactivity was 1.22 ( 0.19, which suggests that
Pgp, this figure shows the results of a single experiment,
the binding of one molecule of colchicine to Pgp is sufficient
but is representative of multiple independent experiments
to cause an increase in the UIC2 reactivity of 3T3-MDR1
demonstrating similar results (not shown). In agreement with
cells. In contrast to colchicine, the Hill number for the
our previous report (10), increasing concentrations of colchi-
increase in the UIC2 reactivity brought about by vinblastine
cine up to 10 mM had no effect on the UIC2 reactivity of
was found to be close to 2 (11). These numbers are in
the K562/i-S9 cell line, derived from human K562 leukemia
agreement with the report that vinblastine has 2 (18, 20) and
cells after retroviral transduction of the human MDR1 gene
colchicine only 1 binding site in Pgp (19).
(b). In contrast, 10 µM vinblastine increased the UIC2
Figure 2 demonstrates that a high concentration (5-10
reactivity of this cell line ∼4-fold (11). Colchicine, however,
mM) of colchicine has differing effects on UIC2 reactivity
induced a 3-fold, dose-dependent increase in UIC2 reactivity
of Pgp in various other multidrug-resistant cell lines express-
in mouse NIH 3T3 cells transduced with a human MDR1-
ing the wild-type human MDR1 Pgp. Colchicine had little
expressing retrovirus [3T3-MDR1 (O)]. To ensure that this
or no effect on the UIC2 reactivity of HeLa-derived HtTA-
effect of colchicine was not due to the ethanol in which
MDR1 cells and of two cell lines derived from the KB-3-1
colchicine was solubilized, we analyzed the effect of
cell line (also a subclone of HeLa) by colchicine selection
increasing ethanol concentrations on the UIC2 reactivity of
(KB-8-5) or by transfection with the wild-type MDR1 cDNA
3T3-MDR1 cells. Ethanol had no effect on the UIC2
(KB-GRC1). Colchicine produced a small increase in the
4326 Biochemistry, Vol. 40, No. 14, 2001
Table 1: Drug Resistance and Maximal Antibody Reactivity Values for Different Cell Lines Expressing the Wild-Type P-Glycoproteinaa The derivations of all cell lines and their drug-sensitive parental lines are described in Experimental Procedures. The LD50 values were determined
from cytotoxicity assays. LD50 values of the drug-resistant cell lines are followed by the LD50 values for the drug-sensitive parental lines in parentheses. The median fluorescence (in arbitrary units) in the presence of saturating concentrations of MRK16 or UIC2 and in the absence or presence of asaturating concentration of either vinblastine or colchicine is listed.
UIC2 reactivity of human MDR1-transfected mouse LMtk-
the absence of vinblastine, was used in place of MRK16
cells (KK-L), a stronger increase in a CEM leukemia cell
reactivity to represent the Pgp levels (data not shown). The
line isolated by multistep selection with vinblastine (CEM/
results depicted in Figure 3A indicate that the Pgp density
VLB-100), and an even stronger increase in the 3T3-FACS14
is the primary determinant of vinblastine resistance in
cell line, derived from NIH 3T3 cells by transduction with
different Pgp-expressing cell lines, in agreement with the
the human MDR1 retrovirus, and in human HT1080 fibro-
previous study, which was limited to the derivatives of a
sarcoma cells transduced with MDR1 (HT1080-MDR1). In
single cell line (20). In contrast to vinblastine resistance,
contrast to the variable effects of colchicine, saturating
however, colchicine resistance showed no correlation with
concentrations of vinblastine consistently increased UIC2
Pgp density (r ) 0.12; Figure 3B), indicating that some
reactivity to a maximal level that was similar to the reactivity
factors in the cellular environment other than the Pgp density
of a conformation-insensitive antibody MRK16 (Table 1) in
affect the ability of Pgp to transport colchicine.
almost all of these cell lines, with the exception of NIH 3T3
We asked if the ability of Pgp to confer colchicine
derivatives where the UIC2 reactivity in the presence of
resistance could be determined by its ability to undergo a
vinblastine remained lower than that of MRK16 (Table 1
conformational transition in the presence of colchicine. If
this were the case, we would expect that relative colchicine
The Ability of Pgp To Confer Colchicine Resistance
resistance, normalized by relative vinblastine resistance (a
Depends on the Cellular EnVironment and Correlates with
function of the Pgp density), would show a Michaelis-
the Effect of Colchicine on UIC2 ReactiVity. We have
Menten dependence on the colchicine-induced increase in
previously shown that the levels of vinblastine resistance in
UIC2 reactivity. Figure 3C shows a type of Lineweaver-
MDR1-transduced NIH 3T3 cell lines correlate with the
Burke plot of the inverse values for the ratio of colchicine
density of Pgp in the cell membrane (20). We asked if similar
resistance to vinblastine resistance relative to the fold increase
correlations could be established for vinblastine and colchi-
in UIC2 reactivity in the presence of saturating amounts of
cine in a comparison of different Pgp-expressing cell types,
colchicine. This plot provides a linear regression with r )
and if the ability of Pgp to confer resistance to different drugs
0.97, indicating a highly significant correlation between the
could be related to the ability of such drugs to increase UIC2
ability of colchicine to induce a UIC2 reactivity shift and
reactivity. For this analysis, we measured the levels of
the relative colchicine resistance conferred by Pgp in different
vinblastine and colchicine resistance in five different drug-
sensitive cell lines and in their multidrug-resistant derivatives,
Colchicine Counteracts the Vinblastine-Induced Increase
expressing the wild-type human MDR1 Pgp. Table 1 shows
in UIC2 ReactiVity. We investigated whether colchicine,
the LD50 values for vinblastine and colchicine resistance for
which does not alter UIC2 reactivity in many of the tested
these multidrug-resistant cell lines and their drug-sensitive
cell lines, would affect the reactivity of such cell lines in
parents (in parentheses). Table 1 also includes the results of
the presence of a saturating concentration of vinblastine.
concurrent FACS assays for immunoreactivity (expressed as
Figure 4A shows the results of such an experiment with the
the mean fluorescence value) of the multidrug-resistant cell
KK-H cell line, which was derived from KK-L cells shown
lines with a conformation-insensitive antibody MRK16
in Figure 2 by selection for increased resistance to vinblas-
specific for the human MDR1 Pgp, with UIC2 in the absence
tine. Cells were reacted with increasing concentrations of
or presence of vinblastine or colchicine, and with the UPC10
colchicine, in the presence or absence of 10 µM vinblastine.
isotype control, the latter reflecting autofluorescence and
While KK-H cells show a strong increase in UIC2 reactivity
nonspecific antibody binding (proportional to the cell
in the presence of vinblastine alone [compare the UIC2
reactivity for the starting point of the KK-H-labeled curve
Using the data in Table 1, we have plotted the relative
with the data point for KK-H cells at the same colchicine
resistance to vinblastine (Figure 3A) or colchicine (Figure
concentration (4) in the absence of vinblastine], their UIC2
3B) against the relative density of Pgp in the individual cell
reactivity is not significantly affected by colchicine alone
lines. This latter parameter was determined by dividing the
(compare the data points in the absence of vinblastine). In
mean MRK16 immunofluorescence by the fluorescence of
the presence of 10 µM vinblastine, however, increasing
cells reacted with UPC10. The data show a strong correlation
concentrations of colchicine steadily reduced the UIC2
between vinblastine resistance and Pgp density, with a
reactivity of KK-H cells, bringing it close to the low level
correlation coefficient (r) of 0.96 (Figure 3A). Similar results
of reactivity observed in the presence of colchicine alone
were obtained when UIC2 reactivity, in the presence or in
(Figure 4A). The best-fit regression using eq 2 in Experi-
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4327
FIGURE 3: Correlations of vinblastine and colchicine resistance in different Pgp-expressing cell lines with Pgp density and UIC2 reactivityshift. Resistance and median fluorescence values for each of the five labeled cell lines are listed in Table 1. The solid line in each panelis the best-fit linear regression, and the error bars represent the standard error of the LD50 (n ) 3) as determined using SigmaPlot. In panelA, relative vinblastine (VLB) resistance (fold increase in LD50 relative to that of the parental cell line) for each Pgp-expressing cell line isplotted on the Y axis vs the ratio of median MRK16 reactivity to median fluorescence of cells stained with the UPC10 control, a measureof Pgp density (X axis). In panel B, the relative colchicine resistance is plotted vs the same measure of Pgp density. Panel C is a type ofLineweaver-Burke plot for the ratio of median UIC2 reactivity to median UIC2 reactivity in the presence of a saturating concentration ofCOL (UIC2+COL) vs the ratio of relative VLB resistance to relative COL resistance.
mental Procedures shows that the Km of colchicine in the
cine (Figure 4B). Surprisingly, both cell lines carrying single
presence of vinblastine in KK-H cells was 6.68 ( 3.01 mM.
NBS mutants of Pgp, KM-H (O) and MK-H (b), demon-
The Hill number was 1.15 ( 0.34, suggesting that 1 molecule
strated a colchicine-dependent increase in UIC2 reactivity.
of colchicine was sufficient to decrease the UIC2 reactivity.
The magnitude of this increase was higher for KM-H than
Essentially the same effects of colchicine in the presence of
for MK-H cells. The solid line through the data in Figure
a saturating concentration of vinblastine were obtained with
4B is the best-fit regression using eq 1 in Experimental
K562/i-S9 cells, except that the Km was 1.96 ( 0.88 mM
Procedures. The Hill numbers for the effect of colchicine
(data not shown). These results indicate that colchicine binds
on KM-H and MK-H cells were close to 1 (0.83 ( 0.50 and
to Pgp and affects its conformation, even in the cells where
1.44 ( 0.25, respectively). Since neither regression line
colchicine alone does not bring about a change in the UIC2
reaches a saturation plateau for colchicine concentration, the
reactivity. Furthermore, we have found that vinblastine, when
apparent affinity of colchicine could not be measured from
tested up to a concentration of 72 µM, is unable to produce
these data. These results indicate that colchicine can increase
a significant increase in the UIC2 reactivity of K562/i-S9
the UIC2 reactivity of Pgp mutants deficient in ATP
cells in the presence of 5 mM colchicine (data not shown).
hydrolysis, even in the cells where it does not alter the
Mutations of the Nucleotide-Binding Sites of Pgp Enhance
reactivity of the functional Pgp. We have previously shown
the Effects of Colchicine on UIC2 ReactiVity. As described
that Pgp mutated in both NBS (MM) has a high UIC2
in the accompanying paper (11), the UIC2 reactivity of Pgp
reactivity which is unchanged in the presence of vinblastine
is affected by nucleotide binding and debinding. We asked
(10). As expected, colchicine also had no effect on the UIC2
if the ability of colchicine to change the UIC2 reactivity
reactivity of the MM mutant (not shown).
would be affected by mutations in the NBS of Pgp. Figure
As with the wild-type KK-H cell line, increasing concen-
4B compares the effects of increasing concentrations of
trations of colchicine decreased the UIC2 reactivity of KM-H
colchicine on UIC2 reactivity of LMtk- cell lines transfected
and MK-H cells in the presence of 10 µM vinblastine to the
with either the wild-type human Pgp (KK-H) or Pgp mutants
levels approaching those that are seen with colchicine alone
carrying K433M or K1076M substitutions of the essential
(Figure 4A). The Hill numbers for this effect of colchicine
lysine residues in the Walker A motifs of the N-terminal
were close to 1 in all three cell lines (1.15 ( 0.34 for KK-
(MK-H) or C-terminal (KM-H) NBS, respectively. These
H, 0.99 ( 0.46 for KM-H, and 1.00 ( 0.15 for MK-H cells).
mutants are capable of binding nucleotides but are devoid
The apparent affinity for this effect of colchicine, however,
of ATPase activity (5), and their UIC2 reactivity is increased
was more than 1 order of magnitude higher for the single
by vinblastine with the same Hill number (2) as the wild-
type Pgp (11). As mentioned above, the UIC2 reactivity of
( 0.04 mM for MK-H cells) than in the wild-type Pgp of
the wild-type Pgp in KK-H cells was unaffected by colchi-
KK-H cells (6.68 ( 3.01 mM). These results provide
4328 Biochemistry, Vol. 40, No. 14, 2001
FIGURE 4: Effects of colchicine on UIC2 reactivity of the wild-type Pgp (A) and of ATPase-deficient Pgp mutants (B) in the presence ofvinblastine. Both panels depict data for LMtk- cells expressing either wild-type Pgp, KK-H, or single-NBS mutants of Pgp, KM-H, andMK-H. In panel A, cells were incubated in the absence [KK-H (3), KM-H (]), and MK-H (0)] or presence of 10 µM vinblastine (b),individually labeled) for 10 min at 37 °C prior to the addition of the indicated concentration of colchicine. The cells were then stained withUIC2 for an additional 30 min at 37 °C. The solid line through each set of data is the best-fit regression as determined using eq 2. In panelB, KK-H (2), KM-H (O), and MK-H (b) cells were stained at the indicated concentration of colchicine for 10 min at 37 °C prior to theaddition of UIC2 and subsequent incubation for an additional 30 min at 37 °C.
additional evidence that mutations that abolish the ATPase
it had in the presence of AMP-PNP. In panel B, colchicine
activity of Pgp increase the effect of colchicine on UIC2
reverses the effect of the nonhydrolyzable analogue almost
as efficiently as vinblastine. Colchicine was similarly effec-
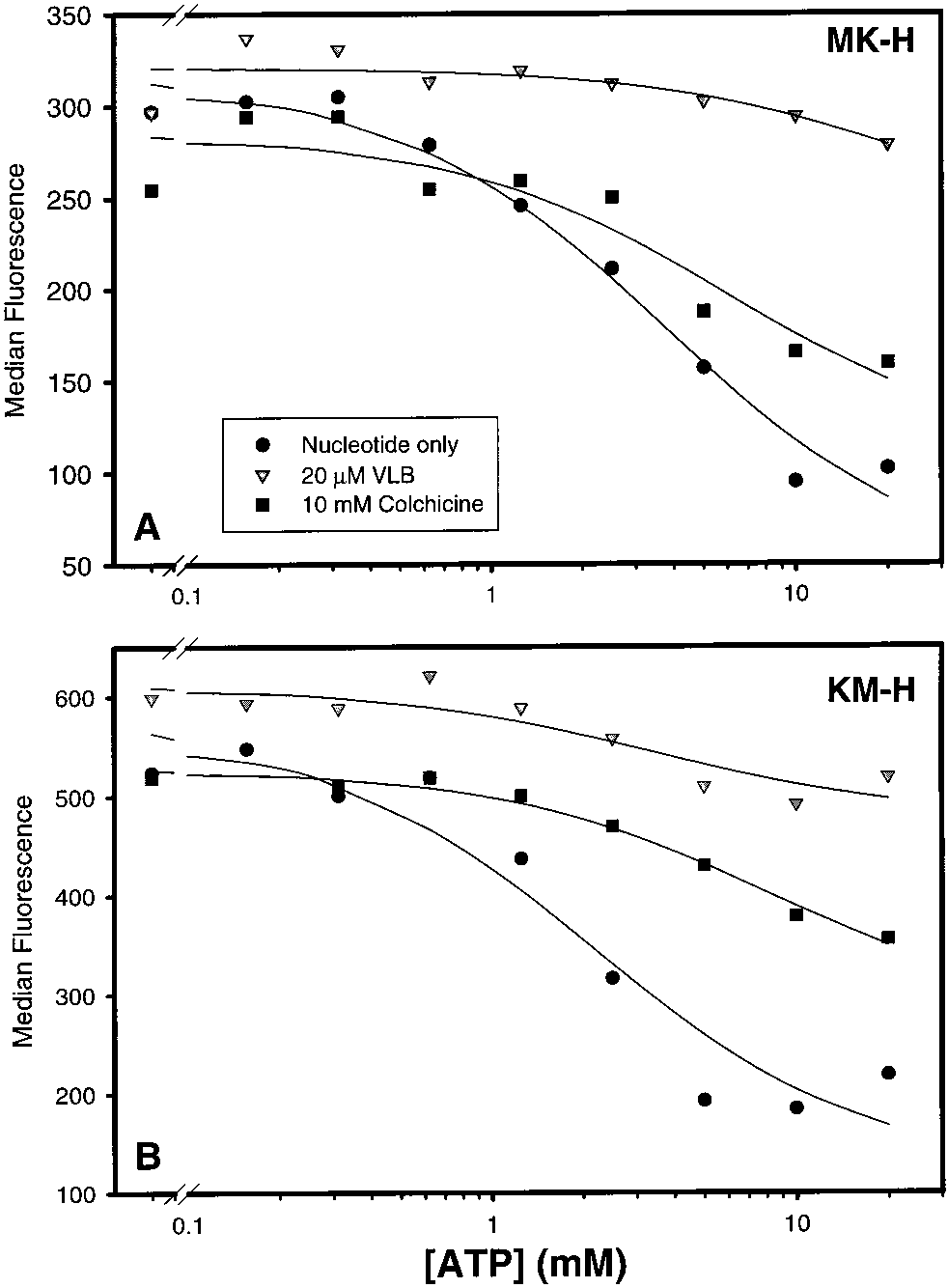
Effects of Different Nucleotides on the UIC2 ReactiVity
tive in reversing the effect of ADP (data not shown). In
in the Presence of Colchicine. To investigate further the
contrast to its effect with nonhydrolyzable nucleotides,
relationship between the conformational effects of colchicine
colchicine provides a moderate increase in UIC2 reactivity
and the nucleotide binding and hydrolysis by Pgp, we
in the presence of lower concentrations of ATP, but at the
analyzed the effects of different nucleotides on the UIC2
highest ATP concentration (20 mM), the UIC2 reactivity is
reactivity in the presence of colchicine, using R-toxin-
unaffected by the presence of colchicine (Figure 5A). An
permeabilized cells. As described in the accompanying paper
important distinction between the decrease in UIC2 reactivity
(11), cell permeabilization depletes cells of endogenous
provided by ATP alone versus ATP and colchicine is found
nucleotide, thereby increasing the UIC2 reactivity to the
when comparing the Hill number for each regression line.
maximal level. The addition of ATP, ADP, or nonhydro-
While the Hill number for ATP alone was approximately 2
lyzable ATP analogues decreases the UIC2 reactivity of
(2.08 ( 0.42), the Hill number for ATP in the presence of
permeabilized cells, but this effect is reversed by the addition
colchicine was 0.60 ( 0.11 (which was not significantly
different from 1). This result suggests that the number of
Figure 5 shows the effects of ATP (panel A) or its
ATP molecules required to decrease UIC2 reactivity is
nonhydrolyzable analogue AMP-PNP (panel B) on the UIC2
decreased in the presence of colchicine.
reactivity of permeabilized KK-H cells in the presence of
Similar experiments were carried out to analyze the UIC2
nucleotide alone (b), nucleotide and 20 µM vinblastine (1),
reactivity of single-mutant Pgps in permeabilized KM-H and
or nucleotide and 10 mM colchicine (9). In agreement with
MK-H cells. As shown in Figure 6, not only vinblastine but
our previous findings (11), both nucleotides provide a dose-
also colchicine was able to reverse the decrease in UIC2
dependent decrease in UIC2 reactivity in the absence of
reactivity provided by ATP. This effect of colchicine in single
drugs, with the Hill number close to 2 for ATP (2.08 ( 0.42;
NBS mutants parallels its ability to increase the UIC2
Figure 5A) and closer to 1 (1.41 ( 0.21; Figure 5B) for
reactivity of intact MK-H and KM-H cells (Figure 4B). As
AMP-PNP. Also as observed in the previous study (11), the
in intact cells, colchicine caused a smaller change in the UIC2
presence of vinblastine (1) reverses the decrease in UIC2
reactivity of MK-H than of KM-H cell line. Colchicine and
reactivity provided by either nucleotide alone (Figure 5A,B).
vinblastine were also effective in reversing the effects of
The addition of the saturating concentration of colchicine,
ADP and AMP-PNP in permeabilized MK-H and KM-H
however, had a different effect in the presence of ATP than
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4329
FIGURE 6: Effects of colchicine on the ATP-induced decrease inUIC2 reactivity in R-toxin-permeabilized MK-H (A) and KM-H
FIGURE 5: Effects of colchicine on the decrease in UIC2 reactivity
of R-toxin-permeabilized cells brought about by ATP (A) or AMP-
PNP (B). Both panels depict KK-H cells that were permeabilized
incubated in the presence of ATP and 10 mM colchicine, and
with the R-toxin prior to the addition of nucleotide and/or drug. In
1) depict data for cells incubated in the presence of ATP
b) depict data for cells incubated in the presence
and 20 µM vinblastine. The solid line through each set of data is
of nucleotide alone (ATP in panel A or AMP-PNP in panel B),
the best-fit regression as determined using eq 2.
squares (9) depict data for cells incubated in the presence ofnucleotide and 10 mM colchicine, and triangles (1) depict datafor cells incubated in the presence of nucleotide and 20 µM
ground depends on the specific Pgp isoforms (21) and on
vinblastine. Cells treated with the R-toxin were incubated in the
the presence of mutations that alter the substrate specificity
presence of nucleotide for 10 min at 37 °C followed by incubation
of Pgp (15). In the study presented here, we asked if relative
for an additional 10 min at 37 °C after the addition of the
resistance to different drugs, conferred by the same wild-
appropriate drug. The solid line through each set of data is the best-
type human MDR1 Pgp, would be affected by the cell type
fit regression as determined using eq 2.
where this Pgp is expressed. We have shown earlier that the
DISCUSSION
resistance to vinblastine, one of the best transport substratesof Pgp, correlated with the cell-surface density of Pgp in
In the study presented here, we compared the ability of
different multidrug-resistant derivatives of the same cell line,
Pgp expressed in different cell types to provide resistance
suggesting that the Pgp level was the principal determinant
to one of its relatively poor transport substrates, colchicine,
of vinblastine resistance in such cells (20). We now extended
and we have correlated this resistance with the ability of
this analysis to compare Pgp-expressing cell lines of different
colchicine to induce conformational transitions of Pgp, which
tissue types, and we still observed the same correlation
can be detected by altered reactivity with a conformation-
between vinblastine resistance and Pgp density. In contrast,
sensitive antibody UIC2. We also compared how colchicine
the relative levels of colchicine resistance varied widely and
affects the Pgp conformation in the presence of another
did not correlate with the Pgp density in different cell types.
transport substrate, vinblastine, in the presence of ATP or
One possible interpretation of this variability was that some
nonhydrolyzable nucleotides, and in the wild-type Pgp
cell-specific mechanisms of colchicine resistance, unrelated
relative to Pgp mutants deficient in ATP hydrolysis. We have
to Pgp, determine the final level of colchicine resistance in
found that the effects of colchicine on Pgp conformation
Pgp-expressing cells. Alternatively, such cell-specific factors
depend on the cellular environment and on the ATP
could still act through Pgp, by influencing its interactions
hydrolysis by Pgp and that these effects correlate with the
with colchicine. Analysis of the effects of colchicine on the
ability of Pgp to confer colchicine resistance in different cell
UIC2 reactivity shift in different cell lines supports the
second interpretation. The levels of colchicine resistance
Previous studies indicated that the relative resistance to
relative to vinblastine resistance in different cell types showed
different Pgp-transported drugs in the same cellular back-
an excellent correlation in a Lineweaver-Burke-type plot
4330 Biochemistry, Vol. 40, No. 14, 2001
with the ability of colchicine to induce UIC2-detectable
also effective in reversing the effect of nonhydrolyzable
conformational transitions. The differences in relative colchi-
nucleotides (AMP-PNP and ADP), suggesting that this
cine resistance or in the effects of colchicine on the Pgp
substrate could also promote nucleotide dissociation. With
conformation cannot be ascribed to the selection history of
ATP, however, colchicine was able to provide partial reversal
the cell lines analyzed in Figure 3. Three of the five lines
of the UIC2 reactivity only at lower ATP concentrations.
were isolated without drug selection after retroviral trans-
Since ATP, but not the nonhydrolyzable nucleotides, was
duction with wild-type human MDR1; one (KB-GRC1) was
able to overcome the conformational effect of colchicine,
derived by transfection with a wild-type MDR1-expressing
this result suggested that ATP hydrolysis could play a role
plasmid vector and a single step of small-scale colchicine
in limiting the ability of colchicine to increase UIC2
selection to isolate the transfectants, and only one line (CEM/
reactivity. This hypothesis was confirmed in the assays using
VLB-100) was derived by multistep selection with vinblas-
the KM-H and MK-H cell lines, which carry mutant forms
tine, which has a potential for selecting additional mutations.
of Pgp that bind, but do not hydrolyze, nucleotide (5). In
The KB-GRC1 and CEM/VLB-100 lines, however, show no
contrast to cells carrying wild-type Pgp, colchicine increased
increase in the resistance to their selective agent relative to
the UIC2 reactivity of intact MK-H and KM-H cells and
the Pgp density, which argues against selection-associated
overcame the effects of the highest ATP concentrations in
mutations in these cell lines. The differences in the colchicine
response in different cell lines can therefore be attributed to
Another important effect of colchicine on the Pgp-ATP
the cellular environment rather than the effects of selection.
interaction is indicated by the finding that the saturating
To the best of our knowledge, this is the first demonstration
concentration of colchicine (10 mM) altered the Hill number
that the cellular environment can alter the relative ability of
for the ability of ATP to decrease UIC2 reactivity from 2 to
Pgp to confer resistance to different drugs.
1. This effect of colchicine may be interpreted in light of a
The finding that colchicine resistance is associated with
recent hypothesis by Sauna and Ambudkar (24). They
colchicine-induced changes in the UIC2 reactivity was
suggest that hydrolysis of 1 ATP molecule is required to
surprising, since colchicine (in contrast to vinblastine and
transport the Pgp-bound substrate, with an associated change
most other Pgp substrates) was previously shown to be
in the Pgp conformation [from E1 to E2, as depicted in the
unable to bring about a change in UIC2 reactivity in K562/
formal scheme given in Figure 6 of the accompanying paper
i-S9 cells (10). Similarly, colchicine was reported to induce
(11)], that lowers substrate affinity. The hydrolysis of a
no change in the proteolytic profile of Pgp, which is another
second ATP molecule would change the Pgp conformation
assay for Pgp conformational transitions (22). By examining
back to E1 to allow the transporter to bind a new substrate
a variety of Pgp-expressing cell lines, we have now found
molecule. Colchicine binding appears to shift the equilibrium
that colchicine increases the UIC2 reactivity in some but not
of these conformational transitions, in the direction from E2
all cell lines. Furthermore, in those lines where colchicine
to E1. In the absence of colchicine, such a shift in equilibrium
alone did not affect UIC2 reactivity, this drug was able to
would otherwise require the hydrolysis of a second ATP
reverse the vinblastine-induced increase in UIC2 reactivity.
molecule. Therefore, in the case of colchicine, only one ATP
This result is consistent with the ability of colchicine to
would be required for the overall conformational transition,
inhibit vinblastine-induced changes in the proteolytic profile
resulting in the decrease in the Hill number for ATP.
of Pgp (22) and with the finding that high concentrations of
What is the nature of the cellular factors that determine
colchicine inhibit vinblastine transport by Pgp (23). Our
the effects of colchicine on the Pgp conformation and the
results suggest that the UIC2 reactivity shift may be used as
ability of Pgp to efflux colchicine? It seems likely that Pgp-
an assay to identify Pgp-interacting agents by their ability
colchicine interactions may be affected by the makeup of
to decrease the UIC2 reactivity in the presence of an
the lipid bilayer in the different cell lines. Several studies
upshifting substrate, such as vinblastine, even if such agents
have reported that changes in the lipid composition of the
by themselves do not alter the UIC2 reactivity.
plasma membrane alter drug and/or nucleotide binding to
In those cell lines where colchicine induces an increase
Pgp. Binding of the substrate [3H]azidopine to reconstituted
in UIC2 reactivity, the increase follows a conventional
Pgp in liposomes was improved when the lipid composition
ligand-binding curve, which can be used to determine the
of the liposomes was increased for cholesterol, stigmasterol,
apparent Km and the Hill number parameters that reflect
or ergosterol, in descending order (25). Also, [3H]azidopine
Pgp-colchicine interactions. In all the cell lines analyzed
photolabeling of Pgp was abolished in the presence of
in the study presented here, the Hill number for colchicine,
nonionic detergents, but not in the presence of urea or a
determined in some lines by the increase in the UIC2
zwitterionic detergent, due presumably to disruption of the
reactivity and in other lines by the decrease in UIC2 reactivity
lipid bilayer (26). Similarly, alterations in the lipid headgroup
in the presence of vinblastine, was approximately 1, sug-
and the acyl chain composition or the lipid bilayer alter the
gesting that the binding of only 1 molecule of colchicine is
apparent affinities of vinblastine, verapamil, and daunorubicin
required to alter the UIC2 reactivity. This finding is
for Pgp, and also affect ATP binding and hydrolysis (9).
consistent with other work reporting that colchicine has a
It is also possible that Pgp interactions may be affected
single binding site on Pgp (19).
by some cytoplasmic factors other than lipid composition.
We have analyzed how the effects of colchicine on UIC2
Thus, Zhang and Ling (27) found that cytoplasmic compo-
reactivity are affected by nucleotide binding and ATP
nents modulate the membrane topology of Pgp molecules
hydrolysis by Pgp. As we have previously shown (11), both
produced in cell-free translation systems, and suggested that
ATP and nonhydrolyzable adenine nucleotides decrease the
Pgp expressed in various cell types may have different
UIC2 reactivity in permeabilized cells, but vinblastine
topological structures. Such topological changes could ac-
efficiently reverses this effect of nucleotides. Colchicine was
count for the differences in proteolytic profiles of Pgp
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4331
observed in the presence of different ligands (22, 28) and
may provide a plausible explanation for altered UIC2
14. Ruth, A. C., Stein, W. D., Rose, E., and Roninson, I. B. (2001)
reactivity. The exact nature of substrate-induced conforma-
tional transitions of Pgp and the cellular factors that affect
15. Choi, K., Chen, C., Kriegler, M., and Roninson, I. (1988) Cell
these transitions remain a subject for future investigation.
16. Chaudhary, P. M., and Roninson, I. B. (1991) Cell 66, 85-
REFERENCES
17. Beck, W. T., Mueller, T. J., and Tanzer, L. R. (1979) Cancer
1. Gottesman, M. M., and Pastan, I. (1993) Annu. ReV. Biochem.
18. van Veen, H. W., Margolles, A., Muller, M., Higgins, C. F.,
2. Ambudkar, S. V., Dey, S., Hrycyna, C. A., Ramachandra, M.,
and Konings, W. N. (2000) EMBO J. 19, 2503-2514.
Pastan, I., and Gottesman, M. M. (1999) Annu. ReV. Phar-
19. Shapiro, A. B., and Ling, V. (1997) Eur. J. Biochem. 250,
3. Higgins, C. F. (1992) Annu. ReV. Cell Biol. 8, 67-113.
20. Choi, K., Frommel, T., Stern, R., Perez, C., Kriegler, M.,
4. Urbatsch, I. L., Sankaran, B., Weber, J., and Senior, A. E.
Tsuruo, T., and Roninson, I. (1991) Proc. Natl. Acad. Sci.
(1995) J. Biol. Chem. 270, 19383-19390.
5. Muller, M., Bakos, E., Welker, E., Varadi, A., Germann, U.
21. Tang-Wai, D. F., Kajiji, S., DiCapua, F., de Graaf, D.,
A., Gottesman, M. M., Morse, B. S., Roninson, I. B., and
Roninson, I. B., and Gros, P. (1995) Biochemistry 34, 32-
Sarkadi, B. (1996) J. Biol. Chem. 271, 1877-1883.
6. Loo, T. W., and Clarke, D. M. (1995) J. Biol. Chem. 270,
22. Wang, G., Pincheira, R., and Zhang, J. T. (1998) Eur. J.
7. Sarkadi, B., Price, E. M., Boucher, R. C., Germann, U. A.,
and Scarborough, G. A. (1992) J. Biol. Chem. 267, 4854-
23. Horio, M., Gottesman, M. M., and Pastan, I. (1988) Proc. Natl.Acad. Sci. U.S.A. 85, 3580-3584.
8. Germann, U. A. (1996) Eur. J. Cancer 32A, 927-944.
24. Sauna, Z. E., and Ambudkar, S. V. (2000) Proc. Natl. Acad.
9. Romsicki, Y., and Sharom, F. J. (1999) Biochemistry 38,
25. Saeki, T., Shimabuku, A. M., Ueda, K., and Komano, T. (1992)
10. Mechetner, E. B., Schott, B., Morse, B. S., Stein, W. D.,
Biochim. Biophys. Acta 1107, 105-110.
Druley, T., Davis, K. A., Tsuruo, T., and Roninson, I. B.
26. Zordan-Nudo, T., Ling, V., Liu, Z., and Georges, E. (1993)
(1997) Proc. Natl. Acad. Sci. U.S.A. 94, 12908-12913.
11. Druley, T. E., Stein, W. D., and Roninson, I. B. (2001)
27. Zhang, J. T., and Ling, V. (1995) Biochemistry 34, 9159-
12. Mechetner, E., and Roninson, I. (1992) Proc. Natl. Acad. Sci.
28. Wang, G., Pincheira, R., Zhang, M., and Zhang, J. T. (1997)
13. Kandel, E. S., Chang, B. D., Schott, B., Shtil, A. A., Gudkov,
A. V., and Roninson, I. B. (1997) Somatic Cell Mol. Genet.
JOB NO. 13-351 C ITY & BO R O U G H O F JU N EA U A LA SK A 'S C A PITA L C ITY CAPITAL CITY FIRE & RESCUE - FIRE March 12, 2013 J O B A N N O U N C E M E N T FIREFIGHTER-EMT I $24.78 Hourly Approximately April 22, 2013 thru September 30, 2013 NOTE: HOURLY RATE BASED ON A 40 HOUR WORK WEEK (3 VACANCIES) Are you looking for summer employment working in the em
Pharmaceutical Care of People with Depression Objectives • Provide an overview of the diagnosis and therapeutic management of depression • Identify key pharmaceutical care needs of this group of patients • Explore ways of positively impacting on the care of this patient population Key aims: • Raise awareness and promoting mental health and well-being • Elimin
 Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4325
FIGURE 1: Effects of different concentrations of colchicine on UIC2reactivity in K562/i-S9 and 3T3-MDR1 cells. Intact K562/i-S9 (b)and 3T3-MDR1 (O) cells were incubated at the indicated concen-trations of colchicine for 10 min at 37 °C prior to the addition ofUIC2 for an additional 30 min at 37 °C.
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4325
FIGURE 1: Effects of different concentrations of colchicine on UIC2reactivity in K562/i-S9 and 3T3-MDR1 cells. Intact K562/i-S9 (b)and 3T3-MDR1 (O) cells were incubated at the indicated concen-trations of colchicine for 10 min at 37 °C prior to the addition ofUIC2 for an additional 30 min at 37 °C.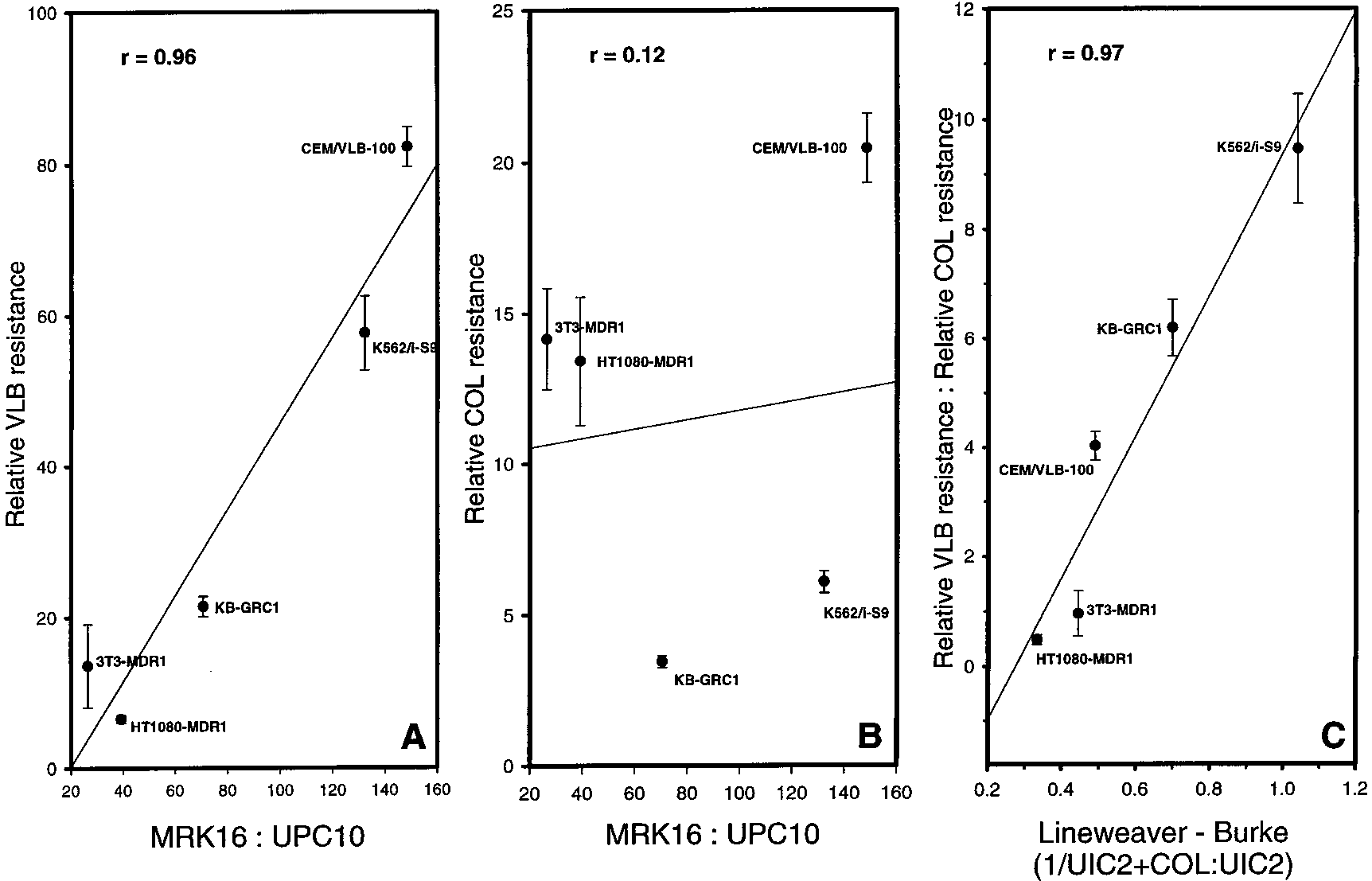 Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4327
FIGURE 3: Correlations of vinblastine and colchicine resistance in different Pgp-expressing cell lines with Pgp density and UIC2 reactivityshift. Resistance and median fluorescence values for each of the five labeled cell lines are listed in Table 1. The solid line in each panelis the best-fit linear regression, and the error bars represent the standard error of the LD50 (n ) 3) as determined using SigmaPlot. In panelA, relative vinblastine (VLB) resistance (fold increase in LD50 relative to that of the parental cell line) for each Pgp-expressing cell line isplotted on the Y axis vs the ratio of median MRK16 reactivity to median fluorescence of cells stained with the UPC10 control, a measureof Pgp density (X axis). In panel B, the relative colchicine resistance is plotted vs the same measure of Pgp density. Panel C is a type ofLineweaver-Burke plot for the ratio of median UIC2 reactivity to median UIC2 reactivity in the presence of a saturating concentration ofCOL (UIC2+COL) vs the ratio of relative VLB resistance to relative COL resistance.
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4327
FIGURE 3: Correlations of vinblastine and colchicine resistance in different Pgp-expressing cell lines with Pgp density and UIC2 reactivityshift. Resistance and median fluorescence values for each of the five labeled cell lines are listed in Table 1. The solid line in each panelis the best-fit linear regression, and the error bars represent the standard error of the LD50 (n ) 3) as determined using SigmaPlot. In panelA, relative vinblastine (VLB) resistance (fold increase in LD50 relative to that of the parental cell line) for each Pgp-expressing cell line isplotted on the Y axis vs the ratio of median MRK16 reactivity to median fluorescence of cells stained with the UPC10 control, a measureof Pgp density (X axis). In panel B, the relative colchicine resistance is plotted vs the same measure of Pgp density. Panel C is a type ofLineweaver-Burke plot for the ratio of median UIC2 reactivity to median UIC2 reactivity in the presence of a saturating concentration ofCOL (UIC2+COL) vs the ratio of relative VLB resistance to relative COL resistance.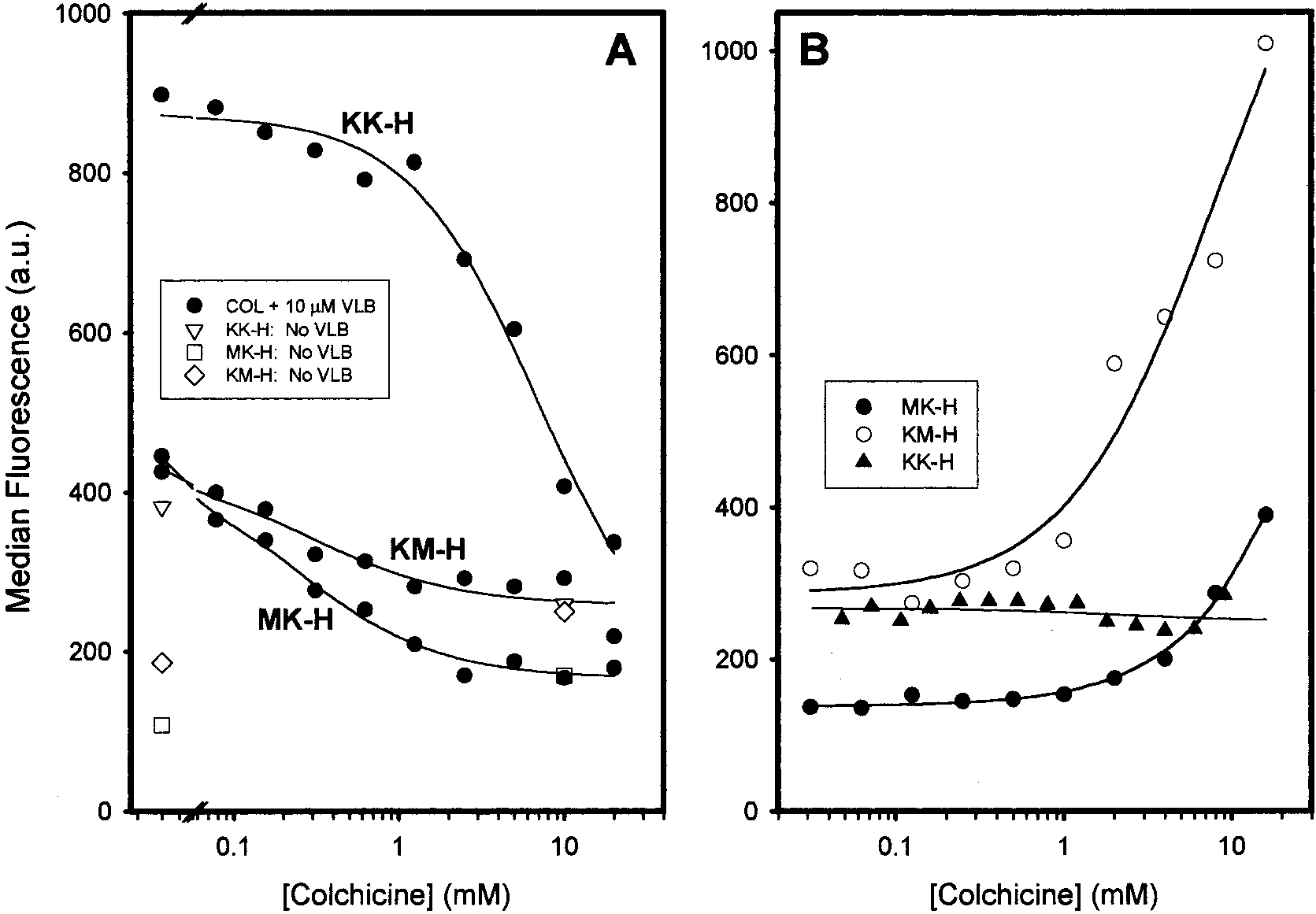 4328 Biochemistry, Vol. 40, No. 14, 2001
FIGURE 4: Effects of colchicine on UIC2 reactivity of the wild-type Pgp (A) and of ATPase-deficient Pgp mutants (B) in the presence ofvinblastine. Both panels depict data for LMtk- cells expressing either wild-type Pgp, KK-H, or single-NBS mutants of Pgp, KM-H, andMK-H. In panel A, cells were incubated in the absence [KK-H (3), KM-H (]), and MK-H (0)] or presence of 10 µM vinblastine (b),individually labeled) for 10 min at 37 °C prior to the addition of the indicated concentration of colchicine. The cells were then stained withUIC2 for an additional 30 min at 37 °C. The solid line through each set of data is the best-fit regression as determined using eq 2. In panelB, KK-H (2), KM-H (O), and MK-H (b) cells were stained at the indicated concentration of colchicine for 10 min at 37 °C prior to theaddition of UIC2 and subsequent incubation for an additional 30 min at 37 °C.
4328 Biochemistry, Vol. 40, No. 14, 2001
FIGURE 4: Effects of colchicine on UIC2 reactivity of the wild-type Pgp (A) and of ATPase-deficient Pgp mutants (B) in the presence ofvinblastine. Both panels depict data for LMtk- cells expressing either wild-type Pgp, KK-H, or single-NBS mutants of Pgp, KM-H, andMK-H. In panel A, cells were incubated in the absence [KK-H (3), KM-H (]), and MK-H (0)] or presence of 10 µM vinblastine (b),individually labeled) for 10 min at 37 °C prior to the addition of the indicated concentration of colchicine. The cells were then stained withUIC2 for an additional 30 min at 37 °C. The solid line through each set of data is the best-fit regression as determined using eq 2. In panelB, KK-H (2), KM-H (O), and MK-H (b) cells were stained at the indicated concentration of colchicine for 10 min at 37 °C prior to theaddition of UIC2 and subsequent incubation for an additional 30 min at 37 °C.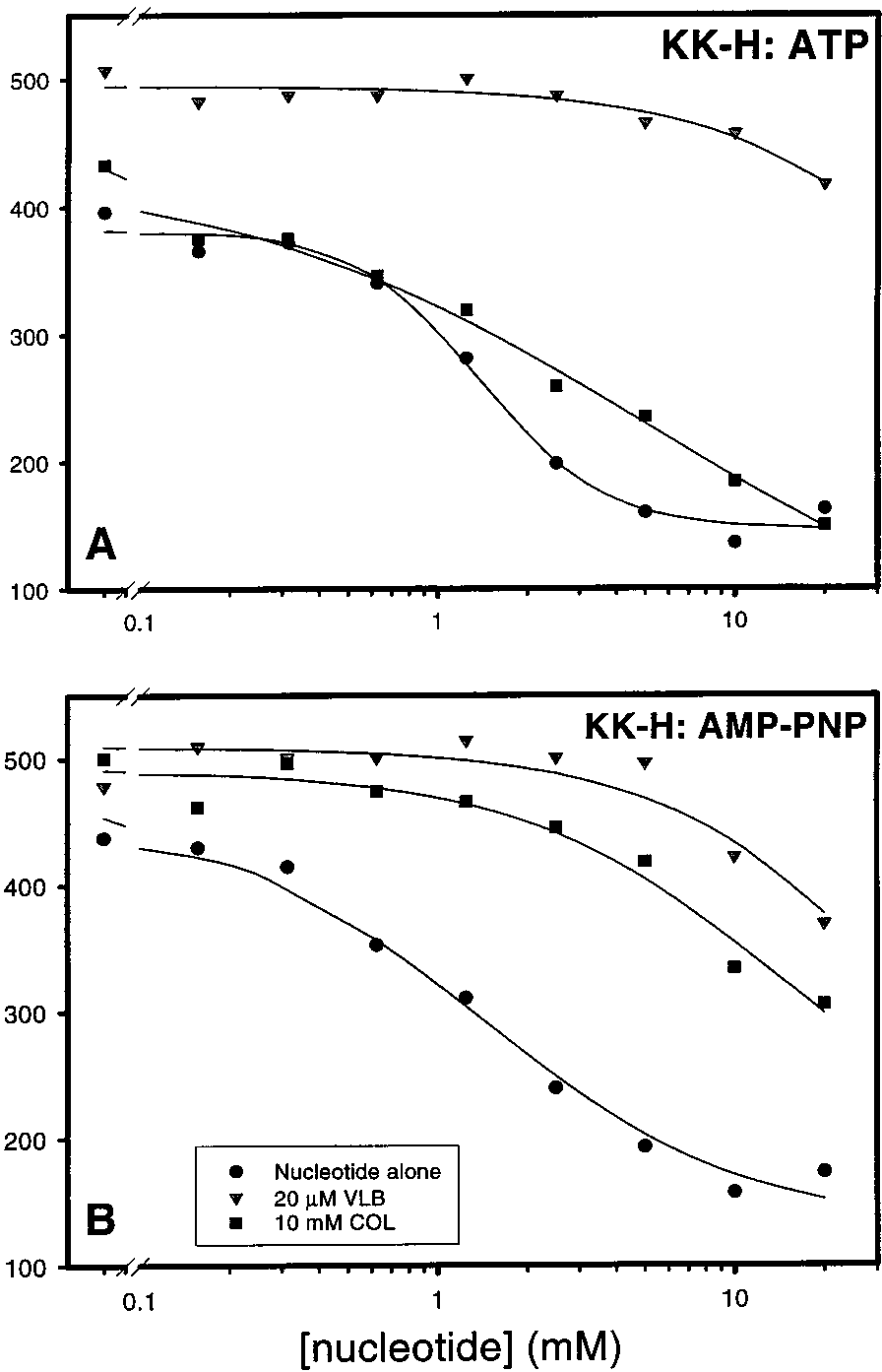
 Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4329
FIGURE 6: Effects of colchicine on the ATP-induced decrease inUIC2 reactivity in R-toxin-permeabilized MK-H (A) and KM-H
FIGURE 5: Effects of colchicine on the decrease in UIC2 reactivity
of R-toxin-permeabilized cells brought about by ATP (A) or AMP-
PNP (B). Both panels depict KK-H cells that were permeabilized
incubated in the presence of ATP and 10 mM colchicine, and
with the R-toxin prior to the addition of nucleotide and/or drug. In
1) depict data for cells incubated in the presence of ATP
b) depict data for cells incubated in the presence
and 20 µM vinblastine. The solid line through each set of data is
of nucleotide alone (ATP in panel A or AMP-PNP in panel B),
the best-fit regression as determined using eq 2.
Colchicine Effects on P-Glycoprotein Conformation
Biochemistry, Vol. 40, No. 14, 2001 4329
FIGURE 6: Effects of colchicine on the ATP-induced decrease inUIC2 reactivity in R-toxin-permeabilized MK-H (A) and KM-H
FIGURE 5: Effects of colchicine on the decrease in UIC2 reactivity
of R-toxin-permeabilized cells brought about by ATP (A) or AMP-
PNP (B). Both panels depict KK-H cells that were permeabilized
incubated in the presence of ATP and 10 mM colchicine, and
with the R-toxin prior to the addition of nucleotide and/or drug. In
1) depict data for cells incubated in the presence of ATP
b) depict data for cells incubated in the presence
and 20 µM vinblastine. The solid line through each set of data is
of nucleotide alone (ATP in panel A or AMP-PNP in panel B),
the best-fit regression as determined using eq 2.