Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Microsoft word - product data sheet2.doc
Species Reactivity: The antibody recognises poly(ADP-ribose) synthesised by a wide
range of poly(ADP-ribose) polymerases like human, mouse, rat or drosophila PARP enzymes.
>95 % as determined by SDS-PAGE. The antibody was purified from bioreactor supernatant using protein A affinity chromatography.
0,1 ml solution containing 50 mM HEPES, 100 mM NaCl, 1% BSA and 0.02 % sodium azide, pH 7.4
Stable for at least 6 months if stored at 2 – 8 °C. upon arrival, store the antibody in suitable aliquots at –20 °C. Avoid repeated freezing and thawing.
For immunofluorescence dilute 10H antibody in 5% nonfat dry milk in PBS to a final concentration of 5-20 µg/ml. For immunodot blot incubate 10H antibody (2,5 µg/ml) in PBS / 0,05% Tween20 / 5% nonfat dry milk.
Monoclonal Antibody (mouse) to Poly(ADP-Ribose) 10H
Monoclonal antibody 10H is directed against poly(ADP-ribose) (PAR) (1). PAR is synthesized after activation of the nuclear DNA repair enzyme poly(ADP-ribose) polymerase (PARP) (4-7). PARP is selectively activated by DNA strand breaks to catalyse the addition of long branched chains of PAR to a variety of nuclear proteins, most notably PARP itself. The amount of PAR formed in living cells with DNA damage is commensurate with the extent of the damage (5). Under DNA damage conditions, PAR undergoes a rapid turnover, with the half-life in the range of minutes, as PAR is rapidly hydrolyzed and converted to free ADP-ribose by the enzyme poly(ADP-ribose)glycohydrolase (PARG). After massive DNA damage (e.g. γ-irradiation or oxidative stress) PAR fluorescence is detectable in the first 10 minutes and disappears later on.
Due to the very large number of endonuclease-mediated DNA breaks in apoptosis, PARP becomes strongly activated during the so-called execution phase. In the case of DNA damage-induced apoptosis, this will represent a “second round” of PAR synthesis. PAR synthesized during apoptosis appears to beremarkably stable. PAR immunofluorescence appears at least as early during apoptosis as does the specific cleavage of PARP by caspase-3. As shown by several groups, this PAR immunofluorescencecorrelates well with other markers of apoptosis (2) (3). References:
Kawamitsu-H; Hoshino-H; Okada-H; Miwa-M; Momoi-H; Sugimura-T; Monoclonal antibodies to poly(adenosine diphosphate ribose) recognize different structures. Biochemistry. 23; 3771-7, (1984)
Negri, C., Donzelli, M., Bernardi, R., Rossi, L., Bürkle, A., Scovassi, A.I. Multiparametric staining to identify apoptotic human cells. Exp. Cell Res., 234: 174-177, (1997).
Bürkle, A., Kretzschmar, H.A., Brown, D.R. Poly(ADP-ribose) immunostaining to detect apoptosis induced by a neurotoxic fragment of prion protein. Histochem. J., im Druck, (1999).
Poly(ADP-Ribose) polymerase and aging Experimental Gerontology, 33: 519-523, (1998)
Bürkle-A; Chen-G; Kupper-JH; Grube-K; Zeller-WJ Increased poly(ADP-ribosyl)ation in intact cells by cisplatin treatment. Carcinogenesis. 14: 559-61 (1993)
Heller, B., Wang, Z.-Q., Wagner, E.F., Radons, J., Bürkle, A., Fehsel, K., Burkart, V., Kolb, H. Inactivation of the poly(ADP-ribose) polymerase gene affects oxygen radical and nitric oxide toxicity in islet cells. J. Biol. Chem., 270, 11176-11180, (1995).
Küpper, J.-H., van Gool, L., Müller, M.,Bürkle, A. Detection of poly(ADP-ribose) polymerase and its reaction product poly(ADP-ribose) by immunocytochemistry. Histochem. J., 28, 391-395, (1996).
Lankenau, S., Bürkle, A., Lankenau, D.-H. Detection of poly(ADP-ribose) synthesis in Drosophila testes upon γ-irradiation. Chromosoma, 108, 44-51, (1999).
HL60 cells (human promyelocytic leukaemia) were induced to undergo apoptosis by treatment with etoposide (67 µM) for 3.5 h. Solvent controls were treated with DMSO (0.1%). After washing of the cells in serum-free medium and adhesion on poly-L-lysine-coated coverslips, cells were fixed in 10 % (w/v) trichloroacetic acid. Washing of the samples in an alcohol, incubation with 10H as first antibody (crude hybridoma supernatant) and with a FITC-conjugated second antibody and the respective washing steps were performed as described in Bürkle et al. (1993). Prior to mounting on slides, samples were incubated with DAPI for 10 min to counterstain cell nuclei. Apoptotic cells, which can be identified in the DAPI staining by chromatin condensation or nuclear fragmentation (1 cell in the control; many cells after etoposide treatment), display intense poly(ADP-ribose)-specific immunofluorescent signals while normal cells show a weak, nonspecific, cytoplasmic background staining. Note that poly(ADP-ribose) formation occuring as an immediate cellular reaction after treatment with oxidants, alkylating agents or ionising radiation is characterised by a fine-granular immunofluorescence pattern visible under high magnification (100x objective) (cf. Bürkle et al., 1993), whereas apoptosis-associated poly(ADP-ribose) formation mostly leads to a homogeneous staining pattern.
Bürkle, A., Chen, G., Küpper, J.-H., Grube, K., Zeller, W.J. (1993) Increased poly(ADP-ribosyl)ation in intact cells by cisplatin treatment. Carcinogenesis (London), 14: 559-561.
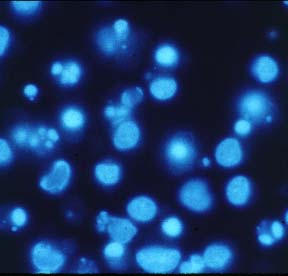
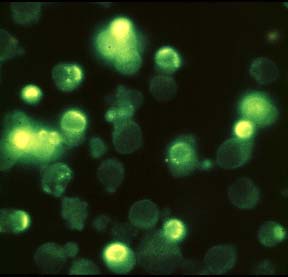
Cells growing on coverslips were incubated with serum free medium, containing 1 mM H2O2 for 10 minutes and then fixed and permeabilized in cool (-20 °C)
methanol/acetone (70:30 v/v) for 10 minutes (-20 °C). Coverslips were air dried and rehydrated in PBS at room temperature for 10 minutes. Samples were than incubated in blocking buffer (PBS, 5% non-fat dry milk, 0.05 % Tween 20) for 10 minutes at room temperature in a humid chamber. Samples were incubated for 1 hr at room temperature with 20µg/ml 10H antibody in blocking buffer. The coverslips were washed five times for 5 minutes each in PBS and incubated with a FITC-conjugated anti-mouse IgG (#F4018, Sigma-Aldrich Chemie GmbH) in blocking buffer containing 1 µg/ml 4,6-diamidino-2-phenylindiole (DAPI). After repeated washing poly(ADP-ribose) was visualized using a fluorescence microscope.
42 antennas for SWLs 1/3/04 11:55 am Page 42If we forget all the theory and come down to hard practice, there are five things aboutan antenna that we can alter. Its length, height, direction, where we feed it and whatsort of wire we use. Graham Brown looks at how we can use this to our advantage. thereof are ‘bad’ is based on diagrams assuming a perfect earth. Ourradiate or receive best at
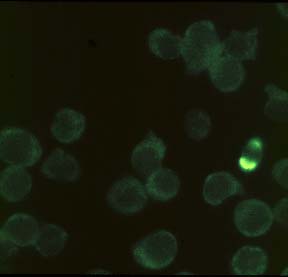
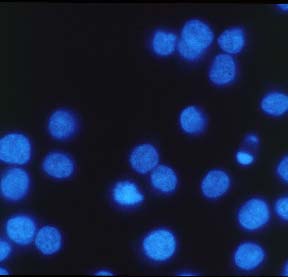
 HL60 cells (human promyelocytic leukaemia) were induced to undergo apoptosis by treatment with etoposide (67 µM) for 3.5 h. Solvent controls were treated with DMSO (0.1%). After washing of the cells in serum-free medium and adhesion on poly-L-lysine-coated coverslips, cells were fixed in 10 % (w/v) trichloroacetic acid. Washing of the samples in an alcohol, incubation with 10H as first antibody (crude hybridoma supernatant) and with a FITC-conjugated second antibody and the respective washing steps were performed as described in Bürkle et al. (1993). Prior to mounting on slides, samples were incubated with DAPI for 10 min to counterstain cell nuclei. Apoptotic cells, which can be identified in the DAPI staining by chromatin condensation or nuclear fragmentation (1 cell in the control; many cells after etoposide treatment), display intense poly(ADP-ribose)-specific immunofluorescent signals while normal cells show a weak, nonspecific, cytoplasmic background staining. Note that poly(ADP-ribose) formation occuring as an immediate cellular reaction after treatment with oxidants, alkylating agents or ionising radiation is characterised by a fine-granular immunofluorescence pattern visible under high magnification (100x objective) (cf. Bürkle et al., 1993), whereas apoptosis-associated poly(ADP-ribose) formation mostly leads to a homogeneous staining pattern.
Bürkle, A., Chen, G., Küpper, J.-H., Grube, K., Zeller, W.J. (1993) Increased poly(ADP-ribosyl)ation in intact cells by cisplatin treatment. Carcinogenesis (London), 14: 559-561.
HL60 cells (human promyelocytic leukaemia) were induced to undergo apoptosis by treatment with etoposide (67 µM) for 3.5 h. Solvent controls were treated with DMSO (0.1%). After washing of the cells in serum-free medium and adhesion on poly-L-lysine-coated coverslips, cells were fixed in 10 % (w/v) trichloroacetic acid. Washing of the samples in an alcohol, incubation with 10H as first antibody (crude hybridoma supernatant) and with a FITC-conjugated second antibody and the respective washing steps were performed as described in Bürkle et al. (1993). Prior to mounting on slides, samples were incubated with DAPI for 10 min to counterstain cell nuclei. Apoptotic cells, which can be identified in the DAPI staining by chromatin condensation or nuclear fragmentation (1 cell in the control; many cells after etoposide treatment), display intense poly(ADP-ribose)-specific immunofluorescent signals while normal cells show a weak, nonspecific, cytoplasmic background staining. Note that poly(ADP-ribose) formation occuring as an immediate cellular reaction after treatment with oxidants, alkylating agents or ionising radiation is characterised by a fine-granular immunofluorescence pattern visible under high magnification (100x objective) (cf. Bürkle et al., 1993), whereas apoptosis-associated poly(ADP-ribose) formation mostly leads to a homogeneous staining pattern.
Bürkle, A., Chen, G., Küpper, J.-H., Grube, K., Zeller, W.J. (1993) Increased poly(ADP-ribosyl)ation in intact cells by cisplatin treatment. Carcinogenesis (London), 14: 559-561.
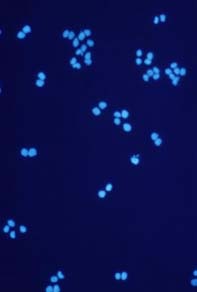
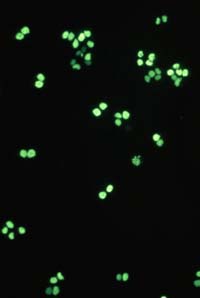
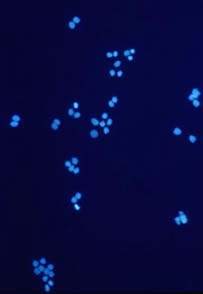
 Cells growing on coverslips were incubated with serum free medium, containing 1 mM H2O2 for 10 minutes and then fixed and permeabilized in cool (-20 °C)
methanol/acetone (70:30 v/v) for 10 minutes (-20 °C). Coverslips were air dried and rehydrated in PBS at room temperature for 10 minutes. Samples were than incubated in blocking buffer (PBS, 5% non-fat dry milk, 0.05 % Tween 20) for 10 minutes at room temperature in a humid chamber. Samples were incubated for 1 hr at room temperature with 20µg/ml 10H antibody in blocking buffer. The coverslips were washed five times for 5 minutes each in PBS and incubated with a FITC-conjugated anti-mouse IgG (#F4018, Sigma-Aldrich Chemie GmbH) in blocking buffer containing 1 µg/ml 4,6-diamidino-2-phenylindiole (DAPI). After repeated washing poly(ADP-ribose) was visualized using a fluorescence microscope.
Cells growing on coverslips were incubated with serum free medium, containing 1 mM H2O2 for 10 minutes and then fixed and permeabilized in cool (-20 °C)
methanol/acetone (70:30 v/v) for 10 minutes (-20 °C). Coverslips were air dried and rehydrated in PBS at room temperature for 10 minutes. Samples were than incubated in blocking buffer (PBS, 5% non-fat dry milk, 0.05 % Tween 20) for 10 minutes at room temperature in a humid chamber. Samples were incubated for 1 hr at room temperature with 20µg/ml 10H antibody in blocking buffer. The coverslips were washed five times for 5 minutes each in PBS and incubated with a FITC-conjugated anti-mouse IgG (#F4018, Sigma-Aldrich Chemie GmbH) in blocking buffer containing 1 µg/ml 4,6-diamidino-2-phenylindiole (DAPI). After repeated washing poly(ADP-ribose) was visualized using a fluorescence microscope.