Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Art. 1.1320
European Review for Medical and Pharmacological Sciences Fluconazole resistance in Candida albicans: a review of mechanisms
I.A. CASALINUOVO, P. DI FRANCESCO, E. GARACI
Department of Experimental Medicine and Biochemical Sciences, Microbiology University of Rome “Tor Vergata” – Rome (Italy)
A b s t r a c t . – Antifungal agents have
Resistance to azole antifungals was reported
greatly contributed to the improvement of pub-
in the late 1980s in C. albicans after prolonged
lic health. Nevertheless, antifungal resistant
therapy with miconazole and ketoconazole. pathogens have increased during the past
Fluconazole is a bis-triazole discovered in
decade, becoming a serious concern. Candida
the 1990s. This compound has been shown to
albicans has been the most extensively stud- ied pathogen in antifungal resistance because
possess potent antifungal activity against
of their morbidity and mortality associated with infections in immunocompromised pa-
such as C. immitis, H. capsulatum, B. dermati-tients. This review describes the antifungal tidis, P. brasiliensis and S. schenckii1. mechanims of the azole fluconazole widely
In spite of its widespread use in the med-
used for the prophylaxis and treatment of can-
ical community, many reports described the
didal infections. The specific molecular path-
clinical failure of fluconazole therapy in indi-
ways occurring in fluconazole-resistance of C. albicans and some issues about new antifun- gal agents are also discussed.
Recently, fluconazole-resistant C. albicans
strains and intrinsically resistant Candida
species such as C. glabrata and C. krusei are
Fluconazole, Candida albicans, Ergosterol, Antifungal
treated for therapy or prophylaxis5-8.
These and other data have led to research
on the molecular mechanisms operating toconfer fluconazole resistance.
In this article we review the current knowl-
edge on the principal resistance mechanisms
Introduction
to fluconazole (Table I). In addition, otherpotential explanations resulting from new ex-
In recent years, fungal infections have in-
perimental data about the above-mentioned
mechanisms are discussed. The findings have
lead to a new therapeutic approach in the
prevention or control of Candida infections.
mune disease and organ or tissue transplan-tion. Candida albicans, a commensal fungusof the oral cavity and gastrointestinal tractin humans, represents one the major causes
Fluconazole
of mucosal infection and systemic infection,which can be life threatening if not treated.
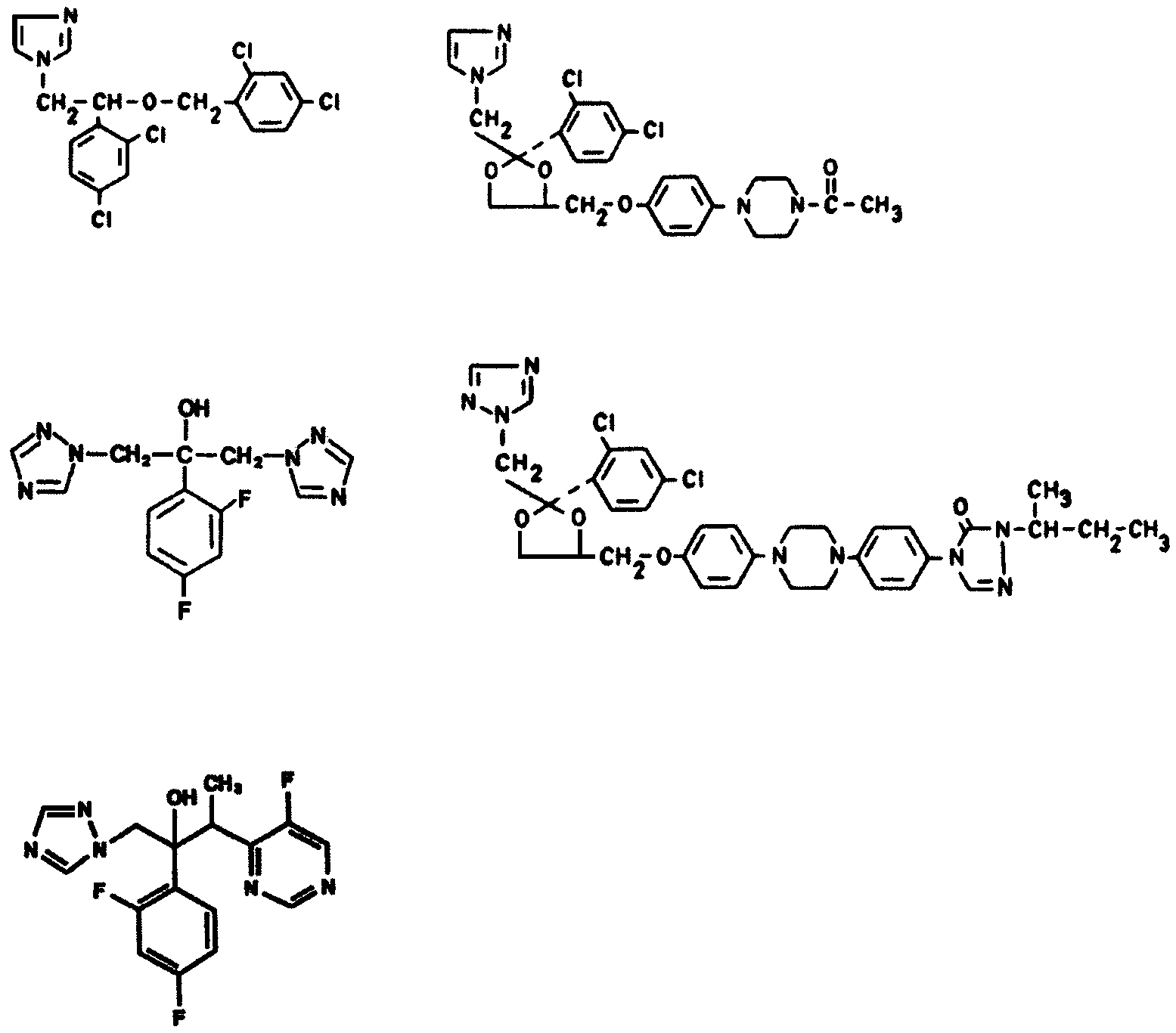
into imidazoles and triazoles (Figure 1).
(azoles, allylamines and morpholines), di-
mazole and ketoconazole) consist of a five-
membered ring structure containing two ni-
(polyenes) or target cell wall biosynthesis
trogen atoms with a complex side chain at-
tached to one of the nitrogen atoms.
I.A. Casalinuovo, P. Di Francesco, E. Garaci
Table I. Overview of fluconazole resistance mechanisms in C. albicans. Molecular basis of Final change accounting References fluconazole resistance for resistance
Reduced drug affinity for the target enzyme
aAllelic differences elimination by gene conversion or mitotic recombination lead to identical mutations in two al-leles; the resulting phenotype is significantly more resistant. bCDRs genes are associated with cross-resistance to other azoles. Miconazole Ketoconazole Fluconazole Itraconazole Voriconazole Figure 1. Chemical structures of azole antifungal agents.
Fluconazole resistance in Candida albicans: a review of mechanisms
Fluconazole and itraconazole are triazole
compounds containing an additional nitrogen
rhoea, hepatotoxicity, have rarely been re-
in the ring9. Other antifungals of new genera-
tion such as posaconazole, ravuconazole and
Prophylactic administration of fluconazole
voriconazole, also belong to triazoles.
has been reserved for selected patients con-
The azole compounds inhibit the lanosterol
sidered to be at high risk of candidemia13. In
demethylase enzyme (or 14α-sterol demethy-
particular, invasive fungal infections have be-
lase); this enzyme converts lanosterol to er-
come increasingly prevalent in individuals
from lanosterol. The 14α-sterol demethylase
neutropenic patients, HIV-infected patients
and transplant recipients. The agreement for
the fluconazole-prophylaxis is still controver-
heme moiety in its active site. The azoles
sial14. However, there is a general consensus
bind to the heme iron through an unhindered
that resistant strains are related to drug expo-
nitrogen, thus inhibiting the enzymatic reac-
tion. In addition, a second nitrogen of the
Fluconazole is fungistatic; this makes it
azoles interacts directly with the apoprotein
clear that host factors contribute to the out-
of lanosterol-demethylase. It is thought that
the affinity of different azoles for the enzymeis also determined by the position of this sec-ond nitrogen10-12. ERG11 and Other ERG Genes
fungal plasma membranes; it is important formembrane integrity and for the activity of
The inhibition of 14α-sterol demethylase
leads to the accumulation of 14α-methylated
demethylase, an essential enzyme for ergos-
sterols, resulting in a defective cell membrane
terol synthesis. Resistance to azole antifungal
with decreased availability of ergosterol and
drugs has been associated with ERG11 gene
overexpression and/or point mutations and
also alterations in the ergosterol biosynthetic
cholesterol. However, the azoles used in ther-
apeutic concentrations demonstrate greater
lanosterol 14α-demethylase and results in in-
affinity for fungal P-450 demethylase than for
the mammalian enzyme. Fluconazole appears
whelms the capacity of the antifungal drug.
to be free of adverse effects on steroid hor-
The effect of ERG11 gene overexpression on
mone production1 and it is available in both
antifungal susceptibility has been described
intravenous and oral formulations. Because
by several studies in C. albicans15-17 and also
of the low toxicity and ready distribution into
in C. glabrata and C. dubliniensis clinical iso-
aqueous body fluids such as cerebrospinal
fluid (CSF), fluconazole has been used in the
treatment of both superficial and systemic
in C. albicans as a consequence of azoles ex-
posure was observed in matched sets of clini-
cal isolates from the same strain20,21. In vitro
discovered in the early 1950s), a favourable
demonstrated in additional Candida species
pharmacokinetic profile (metabolic stability,
such as C. tropicalis, C. glabrata and C.
water solubility) and availability as an oral
and parenteral formulation. These factors
Recently, in an analysis of unmatched sets
have contributed to its therapeutic use in
of clinical isolates it was found that resistance
both normal and immunocompromised hosts.
zole therapy such as nausea, headache, skin
that depletion of the ERG11 gene in C.
I.A. Casalinuovo, P. Di Francesco, E. Garaci
glabrata results in the accumulation of 4,14-
merase was associated with fluconazole resis-
demethylzimosterol, which did not cause de-
tance22,34,35. In contrast, other studies found
fective growth of fungal cells in vitro and in
that the ERG1 gene was repressed in resis-
has also been associated with point mutations
was observed first in S. cerevisiae43. Defective
of the ERG11 gene25-27; these mutations re-
sterol C5,6-desaturase was attributed as the
sult in conformational changes that reduce ef-
cause of fluconazole resistance in C. albicans
fective binding between azoles and their tar-
clinical isolates from AIDS patients44. Such
isolates accumulated ergosterol precursors in-
Several investigators found sequence dif-
ferences of the ERG11 gene in fluconazole-
dienol. The molecular mechanisms associated
resistant C. albicans and in S. cerevisiae
with ERG3 defects are still unclear45,46.
transformants28-30. A list of differentaminoacid exchanges has been provided bydifferent studies that could simply reflect al-lelic variations31. In fluconazole-resistant C.Expression of Two Major albicans isolates frequently observed nu-
Efflux Pumps
cleotide changes were concerned with twoaminoacids located near the heme binding
site (R467K [arginine 467 replaced by lysine]
and G464S [glycine 464 replaced by serine]);
this probably resulted in structural or func-
tional alterations reducing fluconazole affini-
The correlation between decreased suscep-
porters operate through a proton gradient.
tibility to azole drugs and nucleotide changes
CDR2 (Candida Drug Resistance), as well as
that encoding a major facilitator, CaMDR1
Recently, other nucleotide substitutions in
ERG11 gene were identified (K143R [lysine
have been shown to be overexpressed47-51 in
143 replaced by arginine], E266D [glutamic
C. albicans azole-resistant isolates. CaMDR1
acid 266 replaced by aspartic acid], V404L
is specific for fluconazole resistance but not
[valine 404 replaced by leucine], V488I [va-
for other azoles48. Upregulation of these ef-
line 488 replaced by isoleucine]) in three C.
flux pumps reduces the effective concentra-
albicans isolates33; these mutations were asso-
tions of fluconazole in the fungal cell and is
ciated with the fluconazole resistance pheno-
correlated to azole resistance in C. albicans.
type. As suggested by investigators, a single
Genetic deletion of the CDR1 gene resulted
aminoacid change, not interacting with the
in hypersusceptibility to azole drugs52, where-
active site of ERG11p, was unrelated to drug
as CDR2 gene disruption did not cause hy-
resistance. Moreover, mesh membrane struc-
persusceptibility to these agents. The latter
ture developments were observed in the en-
gene is closely related to CDR1 and disrup-
doplasmic reticula of resistant cells33.
Several molecular and genetic studies have
creased hypersusceptibility to azole antifun-
described other ERG genes involved in the
complex ergosterol biosynthesis as alterna-
tive pathways, which were more or less corre-
ed with benomyl resistance in S. cerevisiae)
lated to fluconazole exposure: ERG1, ERG2,
gene deletion in resistant strains of C. albi-cans does not result in increased susceptibili-
In C. albicans, increased expression of
through CDR4) correlated with increased re-
sistance to fluconazole, ketoconazole and
Fluconazole resistance in Candida albicans: a review of mechanisms
itraconazole37. This resistance, however,
Some of the C. albicans cell wall glycopro-
arose rapidly after fluconazole exposure and
teins have been found to be highly immuno-
was transient. In fact, susceptibility resulted
genic and differently modulated according to
in azole-free media and also in vivo after the
drug was no longer administered to the pa-
In vitro studies on the cell wall of flucona-
zole-susceptible and -resistant C. albicans
To date, the molecular mechanisms involv-
strains detected altered distribution of cell
wall glucan-associated proteins63. These re-
CaMDR) have not yet been elucidated.
sults suggest that fluconazole treatment could
Recently, it has been shown that Cdr1p and
have an effect on fungal cell wall metabolism
and structure63,64, and these effects may be
CDR2 genes) in C. albicans act as phospho-
stably incorporated into the cell wall upon ac-
lipids translocators eliciting in-to-out transbi-
The asexual and diploid nature of C. albi-
membrane. It is interesting that fluconazole
cans65,66 complicates the characterization of
could inhibit this transbilayer movement54.
gene expression in antifungal drug resistance.
Several studies investigating changes in chro-
mosome copy number, loss (or not) of het-
Recent results show that in vitro acquired
erozygosity, gene disruption at definite loci
resistance to fluconazole of C. albicans
and other genetic strategies have been linked
strains was associated with variation in mem-
to fluconazole resistance67-70. These studies
brane lipid fluidity and asymmetry56.
show that other factors may contribute to flu-
conazole resistance development. However, a
differences in gene expression identified new
detailed analysis of these and other promising
genes associated or not with drug resistance
in C. albicans. Several of these genes werecoordinately regulated with both CDR genesand CaMDR1, whereas others appeared notto be coordinately regulated with known re-
Different Targets and New
sistance genes35,36. These data suggest that
Therapeutic Approaches
the efflux pumps may be regulated by com-bined expression of several genes. Analysis
of these differentially regulated genes re-
regulatory patterns and new antifungal treat-
quires further investigation and opens up the
ments are currently being undertaken.
possibility of finding new targets for antifun-
pathway and modulation of the susceptibilityto antifungal azoles have been examined. C. albicans mutants in the genes encoding theproteins responsible for cAMP synthesis
Other Changes in Fluconazole Resistance
fluconazole and other sterol biosynthesis in-hibitors71. The addition of cAMP conferred
Recently, antifungal resistance results in
partial-to-complete reversal of this hypersus-
biofilm-associated infections57-59. Efflux
ceptibility. These data suggest that antifungal
pumps do not appear to contribute to flu-
susceptibility could be modulated by adeny-
conazole resistance in C. albicans at late (in-
termediate and mature) stages in biofilm for-
mation60, but solely in the early-phase. On
(CsA) and tacrolimus hydrate (FK506, a 23-
the contrary, changes in sterol profile were
member macrolide) are promising candidates
expressed by resistant phenotypes at interme-
for antifungal therapy, due to their synergis-
diate and mature phases. Therefore, phase-
tic fungicidal effect in combination with
specific mechanisms are suggested to be op-
azoles and non-azole antifungal agents72,73.
erative in antifungal resistance of biofilm
Cyclosporine has several cellular targets in-
I.A. Casalinuovo, P. Di Francesco, E. Garaci
transporters and the cyclophilin-calmodulin-
5) CASE CP, MACGOWAN AP, BROWN NM, REEVES DS,
calcineurin pathway. The mechanism of this
WHITEHEAD P, FELMINGHAM D. Prophylactic oral flu-conazole and candida fungaemia. Lancet 1991;
fungicidal synergism is unknown and was re-
cently reported not to be involved with mul-tidrug efflux transporters74.
6) MARTINS MD, LOZANO-CHIU M, REX JH. Point preva-
lence of oropharyngeal carriage of fluconazole-
resistant Candida in human immunodeficiency
tifungal drugs could also be another thera-
virus-infected patients. Clin Infect Dis 1997; 25:
7) RANGEL-FRAUSTO MS, WIBLIN T, BLUMBERG HM, et al.
are cell wall biosynthesis inhibitors.
National epidemiology of mycoses survey (NE-
MIS): variations in rates of bloodstream infections
spectrum of susceptibility are being proposed
due to Candida species in seven surgical inten-sive care units and six neonatal intensive care
to circumvent cross-resistance within the fun-
units. Clin Infect Dis 1999; 29: 253-258.
In conclusion, in this review we have sum-
RCMERY V, BARNES AJ. Non-albicans Candida spp.
causing fungaemia: pathogenicity and antifungal
marized the mechanisms of resistance to flu-
resistance. J Hosp Infect 2002; 50: 243-260.
conazole in C. albicans. Excellent reviews,
9) BODEY GP. Antifungal agents. In: GP Bodey, ed.
which the reader is referred to, have been
published concerning this matter77-83. Our
Treatment. Raven Press, Ltd, New York: 1993:
10) HITCHCOCK CA. Cytochrome P-450-dependent 14
and/or correlated to antifungal resistance in
alpha-sterol demethylase of Candida albicans
this organism. The resistant phenotype ap-
and its interaction with azole antifungals.
pears to result from different mechanisms
Biochem Soc Trans 1991; 19: 782-787.
not always arising. However it is possible
11) VANDEN BOSSCHE H, MARICHAL P, ODDS FC.
Molecular mechanisms of drug resistance in fun-gi. Trends Microbiol 1994; 2: 393-400.
known. It could be interesting to investigatethe cause of this variability and, if it exists,
12) JOSEPH-HORNE T, HOLLOMON DW. Molecular mecha-
the specific step responsible for fluconazole
resistance. Yet, a combination of different,not only antifungal, drugs could be a promis-
13) EDWARDS JE JR, BODEY GP, BOWDEN RA, et al.
International Conference for the Development of
a Consensus on the Management and Preventionof Severe Candidal Infections. Clin Infect Dis1997; 25: 43-59.
14) SNYDMAN DR. Shifting patterns in the epidemiology
References
of nosocomial Candida infections. Chest 2003;123 (5 Suppl): 500S-503S.
1) GRANT SM, CLISSOLD SP. Fluconazole: A review of
15) LAMB DC, KELLY DE, SCHUNCK WH, et al. The muta-
its pharmacodynamic and pharmacokinetic prop-
tion T315A in Candida albicans sterol 14α-
erties, and therapeutic potential in superficial and
demethylase causes reduced enzyme activity
systemic mycoses. Drugs 1990; 39: 877-916.
and fluconazole resistance through reduced affin-
ity. J Biol Chem 1997; 272: 5682-5688.
Fluconazole-resistant recurrent oral candidiasis in
16) PEREA S, LÓPEZ-RIBOT JL, KIRKPATRICK WR, et al.
human immunodeficiency virus-positive patients:
Prevalence of molecular mechanisms of resis-
persistence of Candida albicans strains with the
tance to azole antifungal agents in Candida albi-
same genotype. J Clin Microbiol 1994; 32: 1115-
cans strains displaying high-level fluconazole re-
sistance isolated from human immunodeficiency
virus-infected patients. Antimicrob Agents
EX JH, RINALDI MG, PFALLER MA. Resistance of
Candida species to fluconazole. Antimicrob
17) LUPETTI A, DANESI R, CAMPA M, DEL TACCA M, KELLY S.
Molecular basis of resistance to azole antifun-
EREA S, LÒPEZ-RIBOT JL, WICKES BL, et al. Molecular
mechanisms of fluconazole resistance in Candida
gals. Trends Mol Med 2002; 8: 76-81.
dubliniensis isolates from human immunodefi-
18) MARICHAL P, VANDEN BOSSCHE H, ODDS FC, et al.
ciency virus-infected patients with oropharyngeal
Molecular biological characterization of an azole-
candidiasis. Antimicrob Agents Chemother 2002;
resistant Candida glabrata isolate. Antimicrob
Agents Chemother 1997; 41: 2229-2237.
Fluconazole resistance in Candida albicans: a review of mechanisms
19) HOLMBERG K, STEVENS DA. Resistance to antifungal
31) MORSCHHÄUSER J. The genetic basis of fluconazole
drugs: current status and clinical implications.
resistance development in Candida albicans.
Curr Opin Anti-Infective Invest Drugs 1999; 1:
Biochim Biophys Acta 2002; 1587: 240-248.
32) MARICHAL P, KOYMANS L, WILLEMSENS S, et al.
20) LÓPEZ-RIBOT JL, MCATEE RK, LEE NL, et al. Distinct
Contribution of mutations in the cytochrome P450
patterns of gene expression associated with de-
14alpha-demethylase (Erg11p, Cyp51p) to azole
velopment of fluconazole resistance in serial
resistance in Candida albicans. Microbiology
Candida albicans isolates from human immuno-
deficiency virus-infected patients with oropharyn-
geal candidiasis. Antimicrob Agents Chemother
Proliferation of intracellular structure correspond-
ing to reduced affinity of fluconazole for cy-
21) WHITE TC, PFALLER MA, RINALDI RG, SMITH J, REDDING
tochrome P-450 in two low-susceptibility strains
SW. Stable azole drug resistance associated with
of Candida albicans isolated from a Japanese
a substrain of Candida albicans from an HIV-in-
AIDS patient. Microbiol Immunol 2003; 47: 117-
fected patient. Oral Dis 1997; 3: S102-S109.
22) HENRY KW, NICKELS JT, EDLIND TD. Upregulation of
34) SMITH WL, EDLIND TD. Histone deacetylase in-
ERG genes in Candida species by azoles and
hibitors enhance Candida albicans sensitivity to
other sterol biosynthesis inhibitors. Antimicrob
azoles and related antifungals: correlation with re-
Agents Chemother 2000; 44: 2693-2700.
duction in CDR and ERG upregulation. AntimicrobAgents Chemother 2002; 46: 3532-3539.
23) WHITE TC, HOLLEMAN S, DY F, MIRELS LF, STEVENS DA.
Resistance mechanisms in clinical isolates of
35) ROGERS PD, BARKER KS. Evaluation of differential
Candida albicans. Antimicrob Agents Chemother
gene expression in fluconazole-susceptible and -
resistant isolates of Candida albicans by cDNA mi-croarray analysis. Antimicrob Agents Chemother
24) NAKAYAMA H, NAKAYAMA N, ARISAWA M, AOKI Y. In vitro
and in vivo effects of 14α-demethylase (ERG11)depletion in Candida glabrata. Antimicrob Agents
36) ROGERS PD, BARKER KS. Genome-wide expression
profile analysis reveals coordinately regulatedgenes associated with stepwise acquisition of
25) WHITE TC. The presence of an R467K aminoacid
azole resistance in Candida albicans clinical iso-
substitution and loss of allelic variation correlate
lates. Antimicrob Agents Chemother 2003; 47:
with an azole-resistant lanosterol 14α-demethy-
lase in Candida albicans. Antimicrob AgentsChemother 1997; 41: 1488-1494.
37) MARR KA, LYONS CN, RUSTAD TR, BOWDEN RA, WHITE
TC. Rapid, transient fluconazole resistance in
26) FRANZ R, KELLY SL, LAMB DC, KELLY DE, RUHNE M,
Candida albicans is associated with increased
MORSCHHÄUSER J. Multiple molecular mechanisms
contribute to a stepwise development of flucona-
zole resistance in clinical Candida albicansstrains. Antimicrob Agents Chemother 1998; 42:
38) DE BACKER MD, ILYINA T, MA XJ, VANDONINCK S, LUYTEN
WH, VANDEN BOSSCHE H. Genomic profiling of theresponse of Candida albicans to itraconazole treat-
27) WHITE TC, MARR KA, BOWDEN RA. Clinical, cellular,
ment using a DNA microarray. Antimicrob Agents
and molecular factors that contribute to antifungal
drug resistance. Clin Microbiol Rev 1998; 11:382-402.
39) BAMMERT GF, FOSTEL JM. Genome-wide expression
patterns in Saccharomyces cerevisiae: compari-
28) SANGLARD D, ISCHER F, KOYMANS L, BILLE J. Amino
son of drug treatments and genetic alterations af-
acid substitutions in the cytochrome P-450
fecting biosynthesis of ergosterol. Antimicrob
lanosterol 14α-demethylase (CYP51A1) from
Agents Chemother 2000; 44: 1255-1265.
azole-resistant Candida albicans clinical isolatescontribute to resistance to azole antifungal
40) MUKHOPADHYAY K, KOHLI A, PRASAD R. Drug suscepti-
agents. Antimicrob Agents Chemother 1998; 42:
bilities of yeast cells are affected by membrane
lipid composition. Antimicrob Agents Chemother2002; 46: 3695-3705.
29) KELLY SL, LAMB DC, KELLY DE. Y132H substitution in
Candida albicans sterol 14alpha-demethylase
41) DIMSTER-DENK D, RINE J, PHILLIPS J, et al. Comprehensive
confers fluconazole resistance by preventing
evaluation of isoprenoid biosynthesis regulation
binding to haem. FEMS Microbiol Lett 1999; 180:
in Saccharomyces cerevisiae utilizing the
Genome Reporter Matrix. J Lipid Res 1999; 40:850-860.
30) LAMB DC, KELLY DE, WHITE TC, KELLY SL. The R467K
amino acid substitution in Candida albicans sterol
42) COWEN LE, NANTEL A, WHITEWAY MS, et al.
14α-demethylase causes drug resistance through
Population genomics of drug resistance in
reduced affinity. Antimicrob Agents Chemother
Candida albicans. Proc Natl Acad Sci U S A
I.A. Casalinuovo, P. Di Francesco, E. Garaci
43) WATSON PF, ROSE ME, ELLIS SW, ENGLAND H, KELLY
55) DOGRA S, KRISHNAMURTHY S, GUPTA V, et al.
SL. Defective sterol C5-6 desaturation and azole
Asymmetric distribution of phosphatidylethanol-
resistance: a new hypothesis for the mode of ac-
amine in C. albicans: possible mediation by
tion of azole antifungals. Biochem Biophys Res
CDR1, a multidrug transporter belonging to ATP
binding cassette (ABC) superfamily. Yeast 1999;15: 111-121.
44) KELLY SL, LAMB DC, KELLY DE, et al. Resistance to
fluconazole and cross-resistance to amphotericin
56) KOHLI A, SMRITI, MUKHOPADHYAY K, RATTAN A, PRASAD R.
B in Candida albicans from AIDS patients caused
In vitro low-level resistance to azoles in Candida
by defective sterol delta5,6-desaturation. FEBS
albicans is associated with changes in membrane
lipid fluidity and asymmetry. Antimicrob AgentsChemother 2002; 46: 1046-1052.
45) JACKSON CJ, LAMB DC, MANNING NJ, KELLY DE, KELLY
SL. Mutations in Saccharomyces cerevisiae sterol
57) CHANDRA J, KUHN DM, MUKHERJEE PK, HOYER LL,
C5-desaturase conferring resistance to the
MCCORMICK T, GHANNOUM MA. Biofilm formation by
CYP51 inhibitor fluconazole. Biochem Biophys
the fungal pathogen Candida albicans: develop-
ment, architecture, and drug resistance. JBacteriol 2001; 183: 5385-5394.
46) SANGLARD D, ISCHER F, PARKINSON T, FALCONER D,
BILLE J. Candida albicans mutations in the ergos-
58) CHANDRA J, MUKHERJEE PK, LEIDICH SD, et al.
terol biosynthetic pathway and resistance to sev-
Antifungal resistance of candidal biofilms formed
eral antifungal agents. Antimicrob Agents
on denture acrylic in vitro. J Dent Res 2001; 80:
47) PRASAD R, DE WERGIFOSSE P, GOFFEAU A, BALZI E.
59) KUHN DM, CHANDRA J, MUKHERJEE PK, GHANNOUM
Molecular cloning and characterization of a novel
MA. Comparison of biofilms formed by Candida
gene of Candida albicans, CDR1, conferring mul-
albicans and Candida parapsilosis on biopros-
tiple resistance to drugs and antifungals. Curr
thetic surfaces. Infect Immun 2002; 70: 878-888.
60) MUKHERJEE PK, CHANDRA J, KUHN DM, GHANNOUM
48) SANGLARD D, KUCHLER K, ISCHER F, PAGANI JL, MONOD
MA. Mechanism of fluconazole resistance in
M, BILLE J. Mechanisms of resistance to azole an-
Candida albicans biofilms: phase-specific role of
tifungal agents in Candida albicans isolates from
efflux pumps and membrane sterols. Infect
AIDS patients involve specific multidrug trans-
porters. Antimicrob Agents Chemother 1995; 39:
PAGNOLI GC, AUSIELLO C, CASALINUOVO I, ANTONELLI
G, DIANZANI F, CASSONE A. Candida albicans and a
49) SANGLARD D, ISCHER F, MONOD M, BILLE J. Cloning of
phosphorylated glucomannan-protein fraction of
Candida albicans genes conferring resistance to
its cell wall induce production of immune interfer-
azole antifungal agents: characterization of
on by human peripheral blood mononuclear cells.
CDR2, a new multidrug ABC transporter gene.
50) FRANZ R, RUHNKE M, MORSCHHÄUSER J. Molecular as-
CASALINUOVO I, CASSONE A. Biochemical and anti-
pects of fluconazole resistance development in
genic characterization of mannoprotein con-
Candida albicans. Mycoses 1999; 42: 453-458.
stituents released from yeast and mycelial formsof Candida albicans. J Med Vet Mycol 1991; 29:
51) WIRSCHING S, MICHEL S, MORSCHHÄUSER J. Targeted
gene disruption in Candida albicans wild-typestrains: the role of the MDR1 gene in fluconazole
63) ANGIOLELLA L, MICOCCI MM, D'ALESSIO S, GIROLAMO A,
resistance of clinical Candida albicans isolates.
MARAS B, CASSONE A. Identification of major glucan-
associated cell wall proteins of Candida albicansand their role in fluconazole resistance. Antimicrob
52) SANGLARD D, ISCHER F, MONOD M, BILLE J.
Agents Chemother 2002; 46: 1688-1694.
Susceptibilities of Candida albicans multidrugtransporter mutants to various antifungal agents
64) HAZEN KC, MANDELL G, COLEMAN E, WU G.
and other metabolic inhibitors. Antimicrob Agents
Influence of fluconazole at subinhibitory concen-
trations on cell surface hydrophobicity andphagocytosis of Candida albicans. FEMS
53) MARR KA, WHITE TC, VAN BURIK JA, BOWDEN RA.
Development of fluconazole resistance inCandida albicans causing disseminated infection
65) PUJOL C, REYNES J, RENAUD F, et al. The yeast
in a patient undergoing marrow transplantation.
Candida albicans has a clonal mode of reproduc-
tion in a population of infected human immunode-ficiency virus-positive patients. Proc Natl Acad
54) SMRITI, KRISHNAMURTHY S, DIXIT BL, GUPTA CM,
MILEWSKI S, PRASAD R. ABC transporters Cdr1p,Cdr2p and Cdr3p of a human pathogen Candida
66) LOCKHART SR, DANIELS KJ, ZHAO R, WESSELS D, SOLL
albicans are general phospholipid translocators.
DR. Cell biology of mating in Candida albicans.
Fluconazole resistance in Candida albicans: a review of mechanisms
67) PEREPNIKHATKA V, FISCHER FJ, NIIMI M, et al. Specific
76) BACHMANN SP, PATTERSON TF, LOPEZ-RIBOT JL. In vitro
chromosome alterations in fluconazole-resistant
activity of caspofungin (MK-0991) against
mutants of Candida albicans. J Bacteriol 1999;
Candida albicans clinical isolates displaying dif-
ferent mechanisms of azole resistance. J ClinMicrobiol 2002; 40: 2228-2230.
68) PUJOL C, MESSER SA, PFALLER M, SOLL DR. Drug re-
sistance is not directly affected by mating type lo-
77) ODDS FC, BROWN AJ, GOW NA. Antifungal agents:
cus zygosity in Candida albicans. Antimicrob
mechanisms of action. Trends Microbiol 2003;
Agents Chemother 2003; 47: 1207-1212.
69) RUSTAD TR, STEVENS DA, PFALLER MA, WHITE TC.
78) GHANNOUM MA, RICE LB. Antifungal agents: mode
Homozygosity at the Candida albicans MTL locus
of action, mechanisms of resistance, and correla-
associated with azole resistance. Microbiology
tion of these mechanisms with bacterial resis-
tance. Clin Microbiol Rev 1999; 12: 501-517.
70) DE BACKER MD, VAN DIJCK P. Progress in functional
79) PEREA S, PATTERSON TF. Antifungal resistance in
genomics approaches to antifungal drug target
pathogenic fungi. Clin Infect Dis 2002; 35: 1073-
discovery. Trends Microbiol 2003; 11: 470-478.
71) JAIN P, AKULA I, EDLIND T. Cyclic AMP signaling
80) SANGLARD D. Clinical relevance of mechanisms of
pathway modulates susceptibility of Candida
antifungal drug resistance in yeasts. Enferm
species and Saccharomyces cerevisiae to anti-
Infecc Microbiol Clin 2002; 20: 462-469.
fungal azoles and other sterol biosynthesis in-
81) COWEN LE, ANDERSON JB, KOHN LM. Evolution of
hibitors. Antimicrob Agents Chemother 2003; 47:
drug resistance in Candida albicans. Annu Rev
72) CRUZ MC, GOLDSTEIN AL, BLANKENSHIP JR, et al.
82) SANGLARD D, ODDS FC. Resistance of Candida
Calcineurin is essential for survival during mem-
species to antifungal agents: molecular mecha-
brane stress in Candida albicans. EMBO J 2002;
nisms and clinical consequences. Lancet Infect
73) ONYEWU C, BLANKENSHIP JR, DEL POETA M, HEITMAN J.
83) LOEFFLER J, STEVENS DA. Antifungal drug resistance.
Ergosterol biosynthesis inhibitors become fungici-
Clin Infect Dis 2003; 36 (Suppl 1): S31-S41.
dal when combined with calcineurin inhibitorsagainst Candida albicans, Candida glabrata, andCandida krusei. Antimicrob Agents Chemother2003; 47: 956-964.
74) MARCHETTI O, MOREILLON P, ENTENZA JM, et al.
Fungicidal synergism of fluconazole and cy-closporine in Candida albicans is not dependent onmultidrug efflux transporters encoded by the CDR1,CDR2, CaMDR1, and FLU1 genes. AntimicrobAgents Chemother 2003; 47: 1565-1570. Acknowledgments
75) DI FRANCESCO P, GAZIANO R, CASALINUOVO IA, et al.
We thank Emanuele Rodolà for his excellent tech-
Combined effect of fluconazole and thymosin al-
nical assistance. This work was supported by MI-
pha 1 on systemic candidiasis in mice immuno-suppressed by morphine treatments. Clin Exp
UR, 60% and progetto FIRB n° RBNE01P4B5-
Antibiotics Improve Survival in Severe Malnutrition Oral antibiotics amoxicillin and cefdinir show efficacy in a randomized study of children with severe acute malnutrition. Linda MacArthur, PhD February 14, 2013—In children with severe acute malnutrition, a one week course of amoxicillin or cefdinir, combined with ready-to-use therapeutic food (RUTF), improved nutritional recovery and
OPEN UNIVERSITY OF MAURITIUS EMPLOYABILITY SKILLS PROGRAMME: (EDUCATION IN NUTRITION) COURSE TITLE: HEALTH AND NUTRITION- [OUES012] LECTURER: DR. VISHNEE BISSONAUTH JUNE 2013 1. Introduction Every day we are faced with an abundance of food choices and nutritional information. Whether to maintain a balanced diet at a restaurant, to browse the aisles of grocery store or
 I.A. Casalinuovo, P. Di Francesco, E. Garaci
Table I. Overview of fluconazole resistance mechanisms in C. albicans.
I.A. Casalinuovo, P. Di Francesco, E. Garaci
Table I. Overview of fluconazole resistance mechanisms in C. albicans.