Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Schwoebel (2001) pain and the body schema. evidence for peripheral effects on mental representations of movement
Brain (2001), 124, 2098–2104
Pain and the body schemaEvidence for peripheral effects on mental representations ofmovement
John Schwoebel,1,2 Robert Friedman,3 Nanci Duda2 and H. Branch Coslett2,1
1Moss Rehabilitation Research Institute, 2Department ofCorrespondence to: H. Branch Coslett, University ofNeurology, University of Pennsylvania School of MedicinePennsylvania Medical Center, 3rd Floor Gates Bldg,and 3Department of Anesthesiology, Temple University3400 Spruce St, Philadelphia, PA 19104-4283, USASchool of Medicine, Philadelphia, Pennsylvania, USAE-mail hbc@mail.med.upenn.eduor Robert Friedman, Department of Anesthesiology,Temple University School of Medicine, 3401 N. Broad St.,Philadelphia, PA 19140, USAE-mail FRIEDMR@tuhs.temple.eduSummary Some accounts of body representations postulate a real- stimulus hand. We found that, as in previous investiga- time representation of the body in space generated by tions, participants’ response times (RTs) reflected the proprioceptive, somatosensory, vestibular and other degree of simulated movement as well as biomechanical sensory inputs; this representation has often been termed constraints of the arm. Importantly, a significant the ‘body schema’. To examine whether the body schema interaction between the magnitude of mental rotation and is influenced by peripheral factors such as pain, we asked limb was observed: RTs were longer for the painful arm patients with chronic unilateral arm pain to determine than for the unaffected arm for large-amplitude imagined the laterality of pictured hands presented at different movements; controls exhibited symmetrical RTs. These orientations. Previous chronometric findings suggest that findings suggest that the body schema is influenced by performance on this task depends on the body schema, pain and that this task may provide an objective measure in that it appears to involve mentally rotating one’s hand from its current position until it is aligned with the Keywords: pain; body schema; plasticity; neglect; parietal Abbreviations: RT ϭ response time, CRPS ϭ complex regional pain syndrome Introduction Take a moment to try the following experiment. With your
Classic neuropsychological observations led Head and
eyes closed, place your left hand in front of you with the
Holmes to postulate an on-line representation of body posture,
palm up and fingers pointed straight ahead. Now rotate your
or ‘body schema’, derived from multimodal sensory inputs
hand until the fingers point first to your right and then to
(including proprioceptive, vestibular, somatosensory and
your left. For the purposes of this paper, there are two
visual inputs) which interacted with motor systems and served
noteworthy observations concerning your ability to perform
to guide movements such as those in the above example
the above tasks. First, the simple fact that such movements
(Head and Holmes, 1911–1912). More recently, several lines
are made effortlessly and without vision suggests that there
of evidence have provided support for the postulated body
must be an on-line mental representation of body posture. That
schema and its role in the guidance of movement. For
is, without real-time information concerning body position, it
example, Cole and Paillard observed striking impairments in
would be impossible to programme such efficient movements.
even routine movements, such as reaching towards an object
Secondly, note that the movements to the right and left were
or maintaining balance while sitting in a chair, for two
not symmetrical: joint constraints on movement probably
patients who were deprived of sensory input as a result of
forced the movement to the left to be slower and more
sensory neuropathy (Cole and Paillard, 1995). Interestingly,
effortful than the movement to the right.
these patients were able to compensate partially for the lack
of on-line information (e.g. proprioceptive) concerning body
they involve lateral rather than medial mental rotations of
posture by constant visual guidance of movements. Such
the hand. These findings, in conjunction with those of Sirigu
effortful compensation helps to highlight the normally
and colleagues (Sirigu et al., 1995, 1996), suggest that both
automatic and seamless interaction between the body schema
actual and mentally simulated movements may depend on
the body schema. Moreover, functional neuroimaging findings
Neuropsychological evidence also suggests that the
suggest that laterality judgements are associated with
monitoring and updating of body position as well as the
activation in motor and parietal areas which substantially
ability to simulate body movements mentally may be impaired
overlap with areas activated by actual movements (e.g.
in patients with parietal damage (Sirigu et al., 1995, 1996;
Stephan et al., 1995; Porro et al., 1996; Parsons and Fox,
Coslett, 1998; Wolpert et al., 1998; Schwoebel et al., 2001).
1998). Parsons and colleagues state ‘In summary, motor
For example, Sirigu and colleagues observed strong
imagery appears generally to involve the same movement
correlations between the time to imagine and execute a series
representation used by the executive motor processes—
of finger movements for both normal subjects and patients
a unitary representation of movements as they occur, in
accordance with the physical laws underlying motor control
movements of the affected limb were both slowed for patients
and implementing all physiological and pathophysiological
with motor cortex damage (Sirigu et al., 1995). However,
constraints.’ (Parsons and Fox, 1998, p. 586).
imagined and executed movement times were poorly
These and other (e.g. Lackner, 1988) lines of evidence
correlated for patients with parietal damage (Sirigu et al.,
suggest that the body schema represents on-line information
1996). Taken together, these data suggest that the parietal
concerning body posture and that it subserves both real and
area is an integral component of the neural substrate for the
imagined movements. Furthermore, the body schema appears
body schema as it appears to be involved in monitoring
to be sensitive to central insults that affect motor performance,
the sensory and motor information necessary for accurate
such as motor cortex lesions and basal ganglia dysfunction
(Dominey et al., 1995; Sirigu et al., 1995). However, to
Parsons further argues that the body schema underlies the
our knowledge, few previous investigations have examined
performance of normal participants on a task that requires
whether there are peripheral factors that influence the
them to judge the laterality of pictured hands (Parsons,
body schema (for a discussion of neural plasticity in
1987a, b, 1994). On the basis of several lines of evidence,
Ramachandran and Hirstein, 1998). The present experiment
judgements by imagining their left hand moving into the
was designed to determine if pain influences the body schema.
orientation of left-hand stimuli and their right hand moving
More specifically, we examined whether performance on a
into the orientation of right-hand stimuli. Furthermore, such
modified version of the hand laterality judgement task
developed by Parsons (Parsons, 1987a) would differ when
representations of the contralateral hand. Consistent with
the judgements involved mental rotations of affected and
these suggestions, Parsons and colleagues (Parsons et al.,
unaffected limbs in patients suffering from chronic arm pain.
1998) found that the accuracy of laterality judgements wasunimpaired in split-brain patients when the stimulus handwas contralateral to the perceiving hemisphere (e.g. a left-hand stimulus presented in the left visual hemifield), but that
performance was not above chance when the stimulus hand
Participants
was ipsilateral to the perceiving hemisphere. Furthermore,
Participants included 13 (six with right arm pain, seven with
Parsons observed that the time required for such laterality
left arm pain) patients with arm pain of at least 3 months
judgements in normal participants increased as the stimulus
duration. Patients were all diagnosed as suffering from
hand was presented at orientations further from that of the
complex regional pain syndrome (CRPS) and were referred
participant’s hand: the time required to judge the laterality
from a pain control centre, where they were undergoing
of a palm-up stimulus hand was modified by whether a
participant’s own hand was palm-up or palm-down and by
characteristics, pain severity and treatment. The medications
the degree of angular disparity between the stimulus hand
prescribed for these patients had a broad range of effects,
and the participant’s hand (Parsons, 1994). Strong correlations
from the pharmacological relief of pain to the anti-seizure
were also observed between the time required for hand
effects of gabapentin and the antidepressant effects of
laterality judgements and the time required for participants
amitriptyline, but we were interested primarily in within-
to actually align their hand with a stimulus. Importantly,
subject comparisons of response times to left- and right-hand
laterality judgement times were also found to reflect
stimuli. Thus, the effects of medications are not likely to
biomechanical constraints on movement. Thus, just as lateral
account for any differences in response times to left- and
movements away from the body’s midline are more effortful
right-hand stimuli within a given patient. Eighteen right-
and time-consuming than medial movements towards the
handed, age-matched (mean age 47 years, SD 11 years)
midline, hand laterality judgement times are also longer when
participants served as controls. Testing was approved by
Table 1 Clinical details of patients with complex regional pain syndrome
Morphine, amitriptyline clonazepam, tramadol
Codeine, gabapentin,rofecoxib, paracetamol
VAS ϭ visual analogue scale: pain severity rated from 0 ϭ no pain to 10 ϭ worst pain experienced. F ϭ female; M ϭ male. Fig. 1 Examples of left-hand stimulus in the palm-down view at orientations of 0°, 90° medial, 90° lateral and 180°.
the Internal Review Board of Temple University and the
Design and procedure
For each patient, the 16 different stimuli (8 conditions ϫ 2
hands) were presented eight times to give a total of 128trials. Controls viewed each stimulus four times in a total of64 trials. Stimuli
Participants sat with their hands resting palm-down on the
Digitized pictures of a left or right hand were presented on
table in front of them with fingers resting on the response
a computer monitor in palm-up and palm-down views at 0°
keys. On each trial, a single stimulus hand appeared centred on
(facing up), 90° medial (facing towards the participant’s
the computer screen and remained there until the participant
midsagittal plane), 90° lateral (facing away from participant’s
indicated the laterality of the hand by pressing a key. For
midsagittal plane) and 180° (facing down) orientations
patients, responses were made by pressing a left or right key
with the index or middle finger of their unaffected limb.
Thus, for both right and left hands there were a total of
Controls responded with the index and middle fingers of
eight different stimuli. All stimuli were created by digitally
their right hand. All participants were instructed to respond
manipulating one picture of a palm-up view and one picture
as quickly and accurately as possible. Psyscope software
of a palm-down view of the same hand in order to ensure
(Cohen et al., 1993) was used to generate a random order
that each stimulus was identical except for the change in
of stimulus presentation for each participant and to record
response time (RT) and accuracy data. Fig. 2 Mean response times for laterality judgements involving Fig. 3 Mean response times for laterality judgements involving
the affected and unaffected limbs of patients for palm-down
the affected and unaffected limbs of patients for palm-up stimuli
stimuli in the four orientation conditions.
[F(1,12) ϭ 7.31, P Ͻ 0.03], indicating slower RTs for mental
A 2 (limb: affected and unaffected arms) ϫ 2 (view: palm
rotations of the affected than the unaffected limb. This effect
up and palm down) ϫ 4 (orientation: 0°, 90° medial, 90°
appears to be driven by slower RTs for the affected limb in
lateral and 180°) repeated measures analysis of variance was
the 180° condition, as indicated by a significant interaction
used to analyse RT and accuracy data separately for patients
between orientation and limb [F(3,36) ϭ 8.12, P Ͻ 0.001].
and controls (for controls, limb refers to the left and right
Planned comparisons yielded significant differences between
arms). Analyses of RT included only data for correct
RTs involving the affected and unaffected limbs for the 180°
responses. RTs Ͼ2 SD above each participant’s grand mean
conditions [palm down, F(1, 12) ϭ 9.58, P Ͻ 0.009; palm
were also excluded from analyses (Ratcliff, 1993), resulting
up, F(1,12) ϭ 7.35, P Ͻ 0.02].
in the loss of 5% of trials for both patients and controls. For
This effect was also consistent across patients. Twelve of
both patients and controls, RT outliers were distributed
the 13 patients exhibited slower RTs for the affected limb
equally across responses involving motor imagery of the left
compared with the unaffected limb for the 180° condition
and right (affected and unaffected) hands, but were more
(sign test, P Ͻ 0.002). On average, mental rotations of the
likely to occur when the disparity between the stimulus hand
affected limb were 1123 ms (SD ϭ 1136 ms) slower than
and the participant’s hand was greatest (i.e. the 180° palm-
those of the unaffected limb in the 180° condition.
Accuracy data for patients and controls are presented in
Table 2. Consistent with the RT data, analyses of accuracyrevealed a significant main effect of orientation [F(3,36) ϭ
Patients
19.29, P Ͻ 0.001] and a significant interaction between
As indicated in Figs 2 and 3, there was a significant main
orientation and view [F(3,36) ϭ 7.07, P Ͻ 0.001], indicating
effect of orientation [F(3,36) ϭ 22.01, P Ͻ 0.001], such that
that accuracy reflected the disparities between stimulus and
RTs increased as the difference between the orientation of
participant hand postures as well as the different constraints
the participant’s hand and the stimulus hand increased. There
on palm-down and palm-up rotations of the hand. There was
was also a significant interaction between orientation and
no significant main effect of limb [F(1,12) Ͻ 1], suggesting
view [F(3,36) ϭ 5.44, P Ͻ 0.005], indicating that RTs
an absence of speed–accuracy trade-offs. Overall accuracy
reflected the disparities between stimulus hands and the
for the affected and unaffected limbs was 84 and 83%,
participant’s own hand postures (i.e. 0° orientation and palm
down) as well as the different movement constraints forpalm-up and palm-down rotations of the hand. Furthermore,consistent with biomechanical constraints on medial and
Controls
lateral movements, RTs were significantly longer for palm-
As indicated in Figs 4 and 5, there was a significant main
up views of 90° lateral compared with 90° medial stimuli
effect of orientation [F(3,51) ϭ 19.95, P Ͻ 0.001] and view
[palm down, F(1,12) ϭ 1.92, P Ͻ 0.19; palm up, F(1,12) ϭ
[F(1,17) ϭ 22.73, P Ͻ 0.001] and an interaction between
orientation and view [F(3,51) ϭ 8.76, P Ͻ 0.001], suggesting
Of greatest import was the significant main effect of limb
that, consistent with the patient data, RTs reflected the
Fig. 4 Mean response times for laterality judgements involving Fig. 5 Mean response times for laterality judgements involving
the left and right limbs of controls for palm-down stimuli in the
the left and right limbs of controls for palm-up stimuli in the four
Table 2 Mean accuracy (proportion correct) for patients
no significant effect of limb. Eleven of 18 controls exhibited
slower RTs for the left than for the right limb for the 180°
condition (sign test, P Ͻ 0.12). RTs for the left limb were,on average, 74 ms slower (SD ϭ 781) than those for the
Consistent with the RT data, analyses of accuracy indicated
a significant main effect of orientation [F(3,51) ϭ 10.13,
Ͻ 0.001] and a significant interaction between orientation
and view [F(3,51) ϭ 7.29, P Ͻ 0.001], indicating that
accuracy reflected the disparities between stimulus and
participant hand positions and the different movement
constraints on palm-up and palm-down rotations of the hand.
There was no significant main effect of limb [F(1,17)
suggesting the absence of speed–accuracy trade-offs. Overall
accuracy for the left and right limbs was 86 and 87%,
Discussion Consistent with previous investigations (Parsons, 1987a, b,
disparities between stimulus and participant hand positions
1994; Parsons and Fox, 1998; Parsons et al., 1998), analyses
and the different movement constraints on palm-up and palm-
for both patients and controls demonstrated that RTs and
down rotations of the hand. Furthermore, there were longer
accuracy were significantly influenced by the degree of
RTs for palm-up views of lateral than medial stimuli [palm
imagined movement necessary to align participants’ hands
down, F(1,17) ϭ 2.19, P Ͻ 0.16; palm up, F(1,17) ϭ 37.03,
with stimuli. Furthermore, RTs were consistent with
P Ͻ 0.001], indicating that RTs reflected the biomechanical
previously observed biomechanical constraints on movement
(e.g. Parsons, 1994). These findings support the suggestion
that the body schema underlies performance on the hand
significance [F(1,17) ϭ 3.83, P Ͻ 0.10], reflecting a small
laterality task. The major, and novel, finding of the present
but consistent advantage for responses involving the right
investigation is that a brain representation of the body is
(i.e. dominant) hand, there was no significant interaction
influenced by pain. Patients, but not controls, exhibited
between orientation and limb [F(3,51) Ͻ 1]. Thus, in contrast
slowed RTs when responses required large-amplitude mental
to the consistently slower RTs observed for the affected
rotations of their affected relative to their unaffected arm.
relative to the unaffected limbs in the 180° condition for the
Before we discuss the theoretical implications of these
patient data (i.e. 12 out of 13), control participants exhibited
data, it is important to emphasize that these findings cannot
be attributed to pain inhibition of movement or ‘guarding’
severity of pain at the time of testing. This hypothesis is
for several reasons. First, patients did not, in fact, move the
painful arm during the experimental task. Nor did patients
The sensory and motor remappings reported in patients
report pain in the course of the imagined movements.
and animals following amputation or deafferentation may
Secondly, the slowing of RTs for the painful arm was
also have an anatomical underpinning that differs from the
observed only in the 180° condition; if guarding were elicited
alteration induced in the body schema by pain. Investigations
automatically by any stimulus depicting the painful extremity,
in animals (Merzenich et al., 1984; Pons et al., 1991) as
one would have expected the slowing of RTs to be observed
well as magnetoencephalogram and transcranial magnetic
for stimuli in all four orientations. We postulate that slowing
stimulation experiments in humans (Cohen et al., 1991;Ramachandran, 1993; Yang et al., 1994; Pascual-Leone et al.,
in the 180° condition occurred because, unlike the other
1996) suggest that the remapping after amputation occurs at
conditions, the 180° condition required large-amplitude
the level of the primary sensory and motor cortices. In
simulated movements at both distal and proximal joints and
contrast, PET investigations of what we term the ‘body
was thus more likely to involve painful regions of the arm
schema’ in humans have suggested that this representation
that tended to include both the elbow and the shoulder.
is supported by posterior parietal and dorsolateral frontal
cortices (Bonda et al., 1996; Parsons and Fox, 1998). As a
demonstrations that pathological conditions may alter the
PET study using the hand laterality task reported in the
body schema. As previously noted, Sirigu and colleagues
present paper demonstrated activation in parietal cortex
reported data from patients with parietal lesions, demonstrat-
(Parsons and Fox, 1998), we believe that the pain-induced
ing that central lesions might disrupt the body schema (Sirigu
alteration in the body schema was likely to be mediated by
et al., 1996). Furthermore, using a task similar to that reported
higher-level sensory cortices of the posterior superior parietal
here, Coslett demonstrated that patients with right-hemisphere
lobes rather than the primary sensory cortex.
lesions resulting in left neglect, but not other patients with
Working with patients suffering from CRPS also called
right hemisphere lesions, exhibited an impaired ability to
reflex sympathetic dystrophy, Galer and colleagues (Galer
identify pictures of left compared with right hands (Coslett,
et al., 1995; Galer and Jensen, 1999; for a similar account,
1998). In the light of previous evidence suggesting that the
see Rommel et al., 1999) have also suggested that the
identification of pictured hands depends on the body schema
frequently observed reduction in movement associated with
(Parsons, 1987a, b, 1994), this asymmetrical performance
this syndrome may be attributable to a central neglect-
suggests that at least some features of the neglect syndrome
like disorder. Their conclusions, however, were based on
may be attributable to disruption of the body schema.
observations of movement and questionnaire data, raising thepossibility that the findings were attributable to guarding. As
The claim that a central representation of the body such
noted previously, the findings from the present experiment,
as the body schema may also be altered by ‘peripheral’
in which participants did not move or report pain, cannot
factors is not without precedent. This phenomenon has been
readily be attributed to this factor.
investigated most extensively in patients with phantom limbs.
In addition to the theoretical implications of the present
As noted by Ramachandran and Hirstein in a recent review,
findings, we note that the hand laterality task, after further
several lines of evidence suggest that, in both animals
examination and modification, may also be of clinical value.
and humans, primary sensory and motor cortices may be
As a blind and objective measure, it may be that performance
‘remapped’ after amputation or deafferentation of a body
on the hand laterality task could provide a more reliable and
part (Ramachandran and Hirstein, 1998). However, the
valid measure of pain than the currently used self-reported
alteration in the body schema exhibited by our patients may
ratings of pain. Furthermore, response time on the hand
differ from that exhibited by patients with phantom limbs.
laterality task may prove to be a more sensitive measure of
Whereas amputation or deafferentation may be expected to
changes in pain than subjective ratings. Finally, we note that
induce long-standing or even permanent changes (but see
the use of the hand laterality task as an assessment tool need
Ramachandran, 1993), the changes in the body schema
not be limited to patients suffering from CRPS, but can be
associated with chronic pain may reflect the current state of
used as an assessment or screening tool for diverse patient
nociceptive (and other sensory) feedback. In this sense, the
populations suffering from motor impairments.
alteration in the body schema exhibited by our patients mayapproximate more closely the distortions of body representa-
Acknowledgements
tions observed when inconsistencies are induced between
We wish to thank Jena Friedman for her valued contribution
multiple sensory inputs (Ramachandran and Hirstein, 1998)
as a research assistant. This work was supported by NIH
and tactile or muscle-stretch inputs (Lackner, 1988). If, as
grant R01 NS37920-02 awarded to H.B.C.
we have argued elsewhere (Coslett, 1998; Buxbaum andCoslett, 2001; Schwoebel et al., 2001), the body schema is
References
an on-line, real-time representation of the position and
Bonda E, Frey S, Petrides M. Evidence for a dorso-medial parietal
possible actions of the body, one might expect the changes
we have observed to be influenced significantly by the
Buxbaum LJ, Coslett HB. Specialised structural descriptions for
lateralized mental representations of hand shape and movement.
human body parts: evidence from autotopagnosia. Cogn Neuro-
Pascual-Leone A, Peris M, Tormos JM, Pascual AP, Catala MD.
Cohen JO, MacWhinney B, Flatt M, Provost J. Psyscope: A
Reorganization of human cortical motor output maps following
new graphic interacting environment for designing psychology
traumatic forearm amputation. Neuroreport 1996; 7: 2068–70.
experiments. Behav Res Meth Instr Comput 1993; 25: 257–71.
Pons TP, Garraghty PE, Ommaya AK, Kaas JH, Taub E, Mishkin M.
Cohen LG, Bandinelli S, Findlay TW, Hallett M. Motor
Massive cortical reorganization after sensory deafferentation in adult
reorganization after upper limb amputation in man. Brain 1991;
macaques. Science 1991; 252: 1857–60.
Porro CA, Francescato MP, Cettolo V, Diamond ME, Baraldi P,
Cole J, Paillard J. Living without touch and peripheral information
Zuiani C, et al. Primary motor and sensory cortex activation during
about body position and movement: studies with deafferented
motor performance and motor imagery: a functional magnetic
subjects. In: Bermudez JL, Marcel A, Eilan N, editors. The body
resonance imaging study. J Neurosci 1996; 16: 7688–98.
and the self. Cambridge (MA): MIT Press; 1995. p. 245–66.
Ramachandran VS. Behavioral and magnetoencephalographic
Coslett HB. Evidence for a disturbance of the body schema in
correlates of plasticity in the adult human brain. [Review]. Proc
neglect. Brain Cogn 1998; 37: 527–44.
Natl Acad Sci USA 1993; 90: 10413–20.
Dominey P, Decety J, Broussolle E, Chazot G, Jeannerod M. Motor
Ramachandran VS, Hirstein W. The perception of phantom limbs:
imagery of a lateralized sequential task is asymmetrically slowed
the D. O. Hebb lecture. [Review]. Brain 1998; 121: 1603–30.
in hemi-Parkinson’s patients. Neuropsychologia 1995; 33: 727–41.
Ratcliff R. Methods for dealing with reaction time outliers. Psychol
Galer BS Jensen M. Neglect-like symptoms in complex regional
pain syndrome: Results of a self-administered survey. J Pain
Rommel O, Gehling M, Dertwinkel R, Witscher K, Zenz M, Malin
J-P, et al. Hemisensory impairment in patients with complex regional
Galer BS, Butler S, Jensen MP. Case reports and hypothesis: a
pain syndrome. Pain 1999; 80: 95–101.
neglect-like syndrome may be responsible for the motor disturbance
Schwoebel J, Coslett HB, Buxbaum LJ. Compensatory coding of
in reflex sympathetic dystrophy (complex regional pain syndrome-1).
body part location in autotopagnosia: evidence for extrinsic
[Review]. J Pain Symptom Manage 1995; 10: 385–91.
egocentric coding. Cogn Neuropsychol 2001; 18: 363–81.
Head H, Holmes G. Sensory disturbances from cerebral lesions.
Sirigu A, Cohen L, Duhamel JR, Pillon B, Dubois B, Agid Y.
Congruent unilateral impairments for real and imagined hand
Lackner JR. Some proprioceptive influences on the perceptual
movements. Neuroreport 1995; 6: 997–1001.
representation of body shape and orientation. Brain 1988; 111:
Sirigu A, Duhamel JR, Cohen L, Pillon B, Dubois B, Agid Y. The
mental representation of hand movements after parietal cortex
damage. Science 1996; 273: 1564–8.
Schoppmann A, Zook JM. Somatosensory cortical map changes
Stephan KM, Fink GR, Passingham RE, Silbersweig D, Ceballos-
following digit amputation in adult monkeys. J Comp Neurol 1984;
Baumann AO, Frith CD, et al. Functional anatomy of the mental
representation of upper extremity movements in healthy subjects.
Parsons LM. Imagined spatial transformations of one’s hands and
feet. Cogn Psychol 1987a; 19: 178–241.
Wolpert DM, Goodbody SJ, Husain M. Maintaining internal
Parsons LM. Imagined spatial transformation of one’s body. J Exp
representations: the role of the human superior parietal lobe. Nat
Parsons LM. Temporal and kinematic properties of motor behavior
Yang TT, Gallen CC, Ramachandran VS, Cobb S, Schwartz BJ,
reflected in mentally simulated action. J Exp Psychol Hum Percept
Bloom FE. Noninvasive detection of cerebral plasticity in adult
human somatosensory cortex. Neuroreport 1994; 5: 701–4.
Parsons LM, Fox PT. The neural basis of implicit movements usedin recognising hand shape. Cogn Neuropsychol 1998; 15: 583–615. Received January 25, 2001. Revised May 17, 2001.
Parsons LM, Gabrieli JD, Phelps EA, Gazzaniga MS. Cerebrally
Personne-ressource: FICHE TECHNIQUE Shampooing et revitalisant Huile d’amande et avocat pour cheveux secs/endommagés Naturals Nom du produit: Shampooing et revitalisant Huile d’amande et avocat pour cheveux secs/endommagés Naturals Description: • Aide à régénérer les cheveux secs, • Aide à redonner aux cheveux une texture Shampooing hydratant
CASE STUDY Functional upper airway obstruction and chronic irritation of the larynx Functional upper airway obstruction and chronic irritation of the larynx. T.B. Rothe, Luzerner Hoehenklinik, Montana, Switzerland. W. Karrer. ERS Journals Ltd 1998. ABSTRACT: Wheezing and dyspnoea are typical symptoms of asthma but can also be found in diseases of the extrathoracic airways.
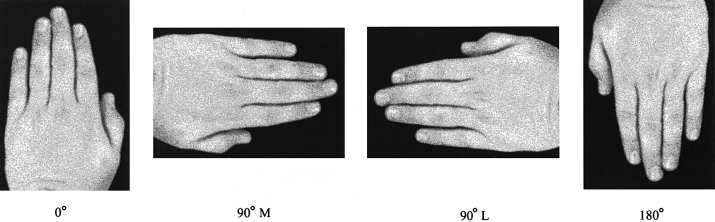 Table 1 Clinical details of patients with complex regional pain syndrome
Table 1 Clinical details of patients with complex regional pain syndrome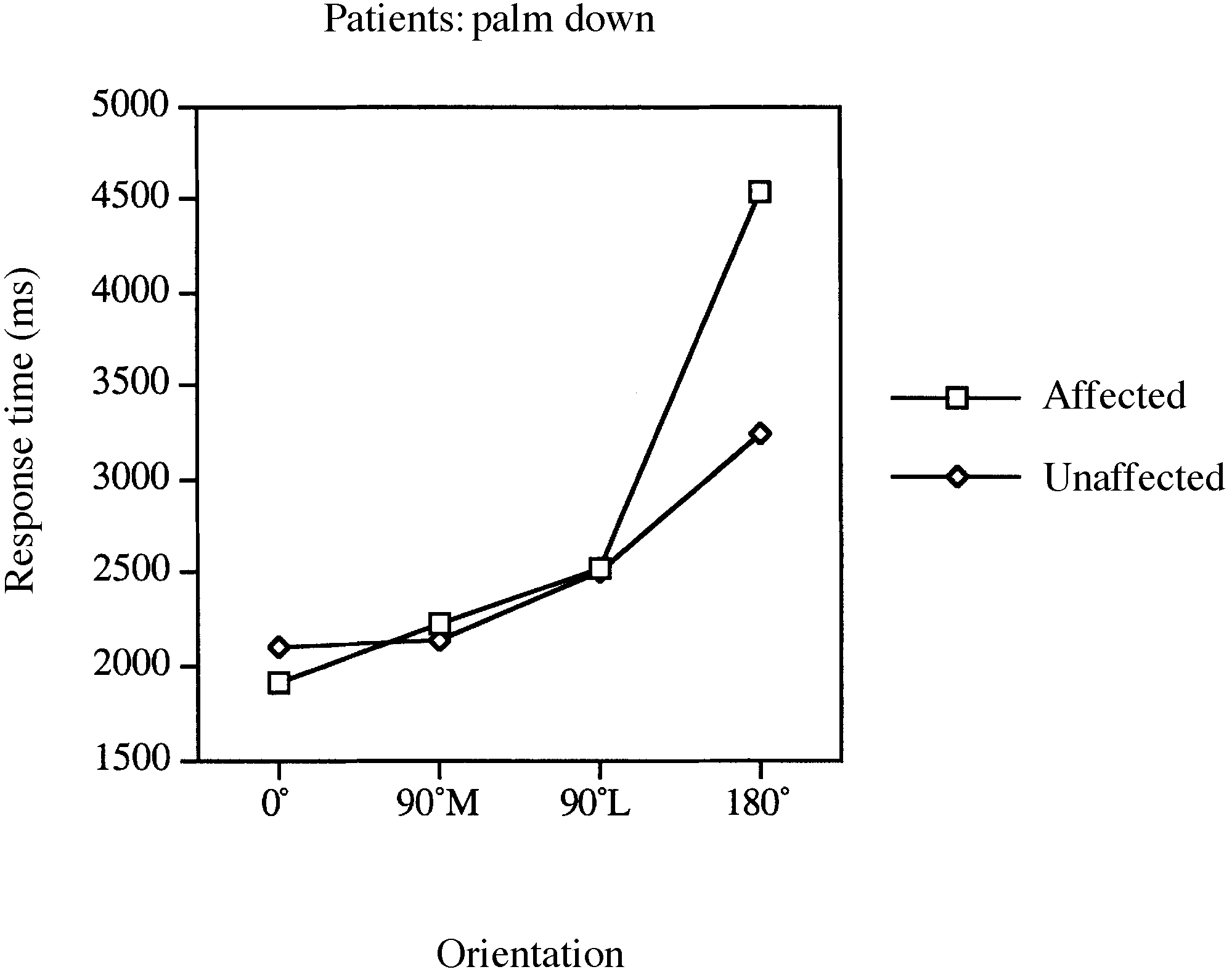
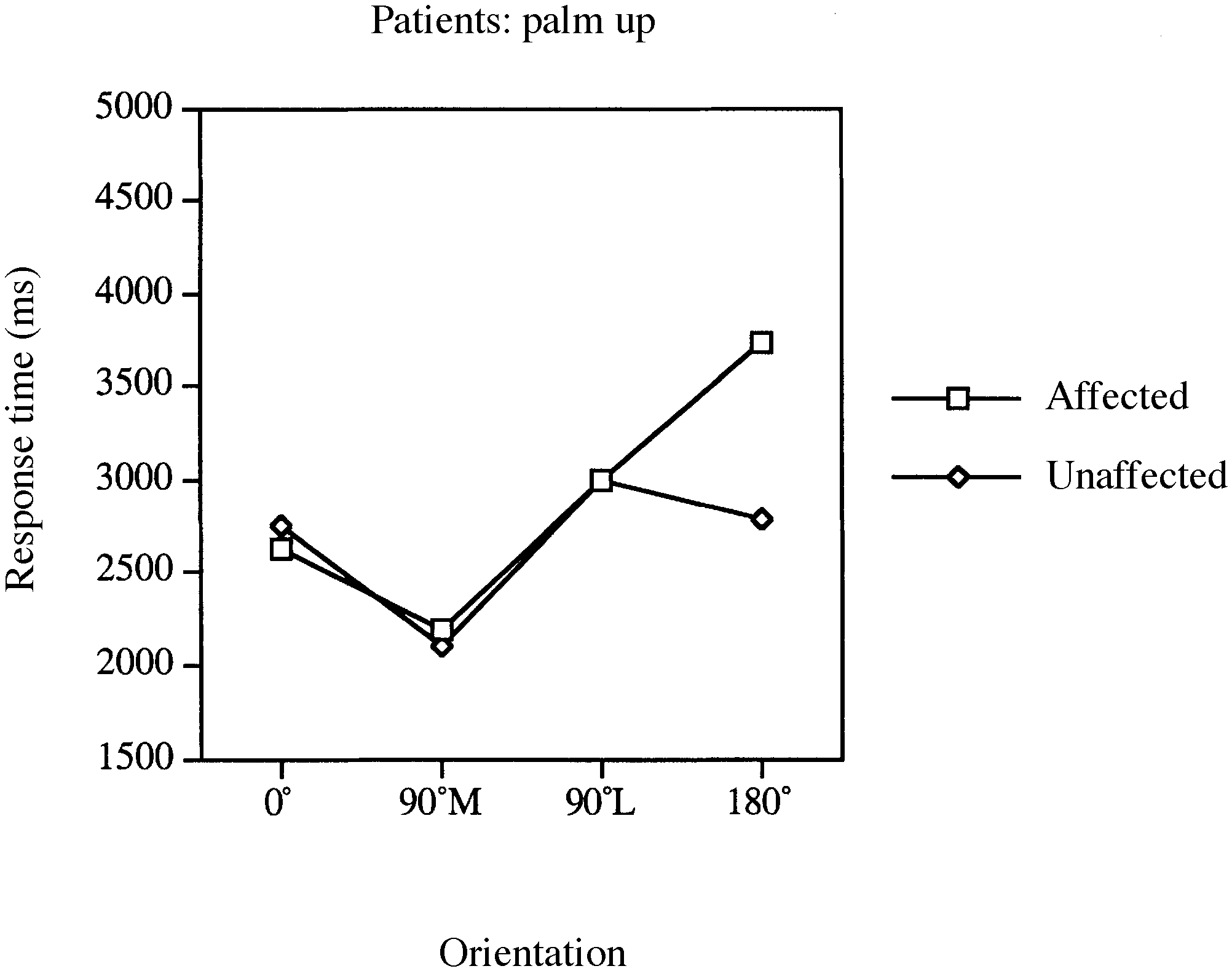 Fig. 2 Mean response times for laterality judgements involving
Fig. 2 Mean response times for laterality judgements involving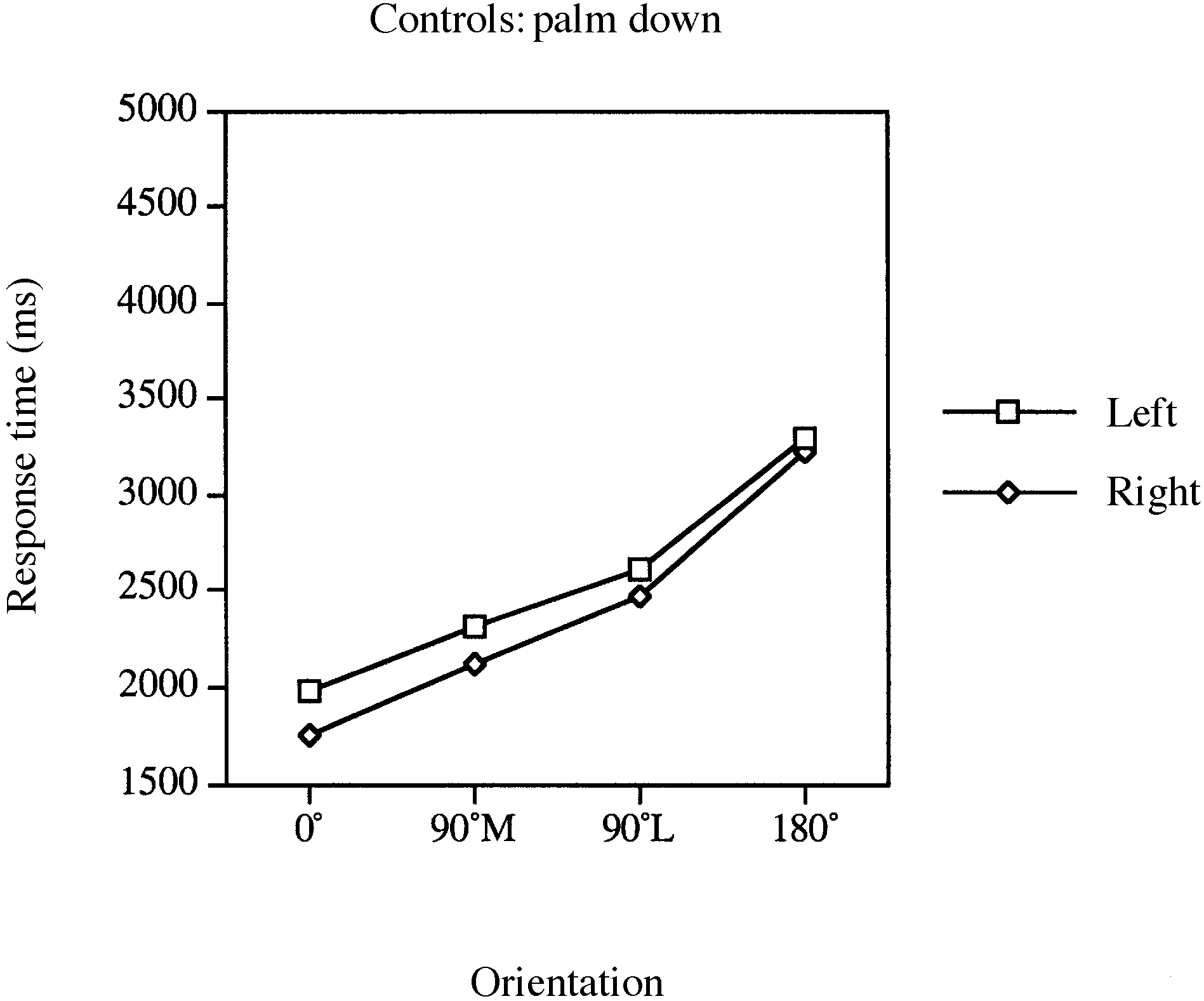
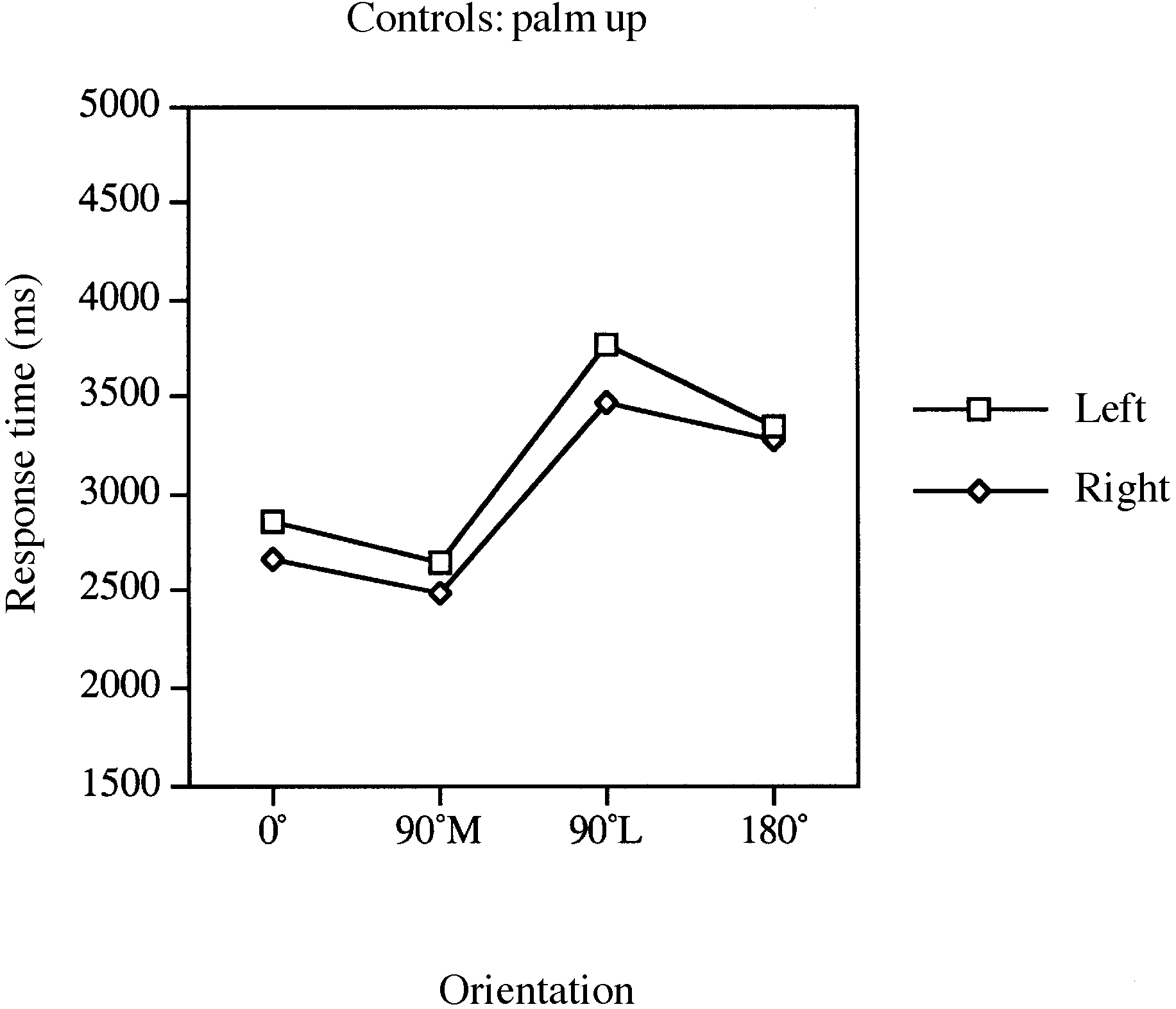 Fig. 4 Mean response times for laterality judgements involving
Fig. 4 Mean response times for laterality judgements involving