Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Brs.kyushu-u.ac.jp
Oncogene (2007), 1–5& 2007 Nature Publishing Group All rights reserved 0950-9232/07 $30.00
p21Waf1/Cip1/Sdi1 mediates retinoblastoma protein degradation
EV Broude1, ME Swift2, C Vivo1, B-D Chang1, BM Davis1, S Kalurupalle1, MV Blagosklonny1and IB Roninson1
1Cancer Center, Ordway Research Institute, Albany, NY, USA and 2Department of Molecular Genetics, University of Illinois atChicago, Chicago, IL, USA
Damage-induced G1 checkpoint in mammalian cells
the proteasome. Proteasome-mediated Rb degradation
involves upregulation of p53, which activates transcription
is promoted by Mdm2 (Sdek et al., 2005) and gankyrin
of p21Waf1 (CDKN1A). Inhibition of cyclin-dependent
(Higashitsuji et al., 2000), E7 of papilloma virus (Boyer
kinase (CDK)2 and CDK4/6 by p21 leads to depho-
et al., 1996) and Tax of HTLV1 (Kehn et al., 2005).
sphorylation and activation of Rb. We now show that
Oncoprotein-induced proteasomal degradation of Rb is
ectopic p21 expression in human HT1080 fibrosarcoma
one of the mechanisms for Rb inactivation in carcino-
cells causes not only dephosphorylation but also depletion
genesis (Ying and Xiao, 2006), but Rb degradation has
of Rb; this effect was p53-independent and susceptible to a
not been described in DNA damage response.
proteasome inhibitor. CDK inhibitor p27 (CDKN1B) also
Changes in Rb phosphorylation are most commonly
caused Rb dephosphorylation and depletion, but another
detected by immunoblotting through changes in the
CDK inhibitor p16 (CDKN2A) induced only depho-
protein’s electrophoretic mobility. Examination of
sphorylation but not depletion of Rb. Rb depletion was
numerous Rb immunoblots published by different
observed in both HT1080 and HCT116 colon carcinoma
groups showed that in many (but not all) cases Rb
cells, where p21 was induced by DNA-damaging agents.
dephosphorylation, which results from drug treatment,
Rb depletion after DNA damage did not occur in the
cell senescence or ectopic p21 expression, is associated
absence of p21, and it was reduced when p21 induction was
with a reduction in the Rb protein signal. In the present
inhibited by p21-targeting short hairpin RNA or by a
study, we have asked (i) whether a decrease in the Rb
transdominant inhibitor of p53. These results indicate that
signal in response to p21 reflects protein degradation or
p21 both activates Rb through dephosphorylation and
merely altered immunoreactivity of dephosphorylated
inactivates it through degradation, suggesting negative
Rb, (ii) if p53 plays a p21-independent role in the de-
feedback regulation of damage-induced cell-cycle check-
crease in Rb, (iii) whether such decrease can be induced
by other CDK inhibitors that induce Rb dephosphory-
Oncogene advance online publication, 7 May 2007;
lation and (iv) if the decrease in Rb in drug-damaged
cells is dependent on p21 induction.
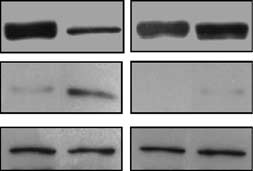
We have described previously a subline of HT1080
cells, p21-9, which expresses p21 from a promoterinducible by b-galactoside isopropyl-b-thio-galactoside(IPTG) (Chang et al., 1999). p21 induction in HT1080p21-9 leads to Rb dephosphorylation and depletion
p53-inducible cyclin-dependent kinase (CDK) inhibitor
without any changes in Rb mRNA levels (Chang et al.,
p21 (CDKN1A) is the key mediator of damage-induced
2000). As shown in Figure 1a (left lanes), Rb was
cell-cycle arrest. p21 interacts with different cyclin/CDK
dephosphorylated and its signal intensity decreased 24–
complexes and other regulators of transcription and
48 h after IPTG addition. To determine if the reduced
signal transduction, exerting broad effects on cell
Rb signal indicates a decrease in protein levels rather
survival, gene expression and morphology (Roninson,
than altered antibody reactivity, we have used both a
2002). p21 effects are partially mediated by Rb, which is
mouse monoclonal and a rabbit polyclonal antibody
inactivated in proliferating cells through phosphoryla-
against Rb. Both antibodies produced the same result
tion by CDK2 and CDK4/6, both of which are inhibi-
(Figure 1a, left lanes), indicating that the decrease in Rb
ted by p21. As a result, p21 induction leads to Rb
signal intensity was indeed due to a decrease in the
dephosphorylation and activation, with ensuing G1
The decrease in Rb protein was not accompanied by
Whereas p21 activates Rb by dephosphorylation,
the appearance of Rb fragments that arise after caspase
several oncoproteins inactivate Rb by degradation via
cleavage of Rb (Tan et al., 1997). Since the best-knownmechanism of Rb degradation is via the proteasomepathway (Ying and Xiao, 2006), we have tested the
Correspondence: Dr IB Roninson, Cancer Center, Ordway Research
effect of a proteasome inhibitor ALLN (N-acetyl-
Institute, 150 New Scotland Avenue, Albany, NY 12208, USA.
leucinyl-leucinyl-norleucinal) on p21-induced Rb deple-
E-mail: roninson@ordwayresearch.orgReceived 21 February 2007; accepted 18 March 2007
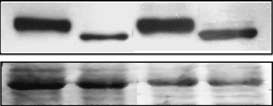
tion. In the experiment shown in Figure 1b, cells were
treated with IPTG for 20 h with ALLN added in one
likely reflects the effect of GSE56 on the p53 response
arm of the experiment for the last 4 h (longer treatment
element present in the cytomegalovirus (CMV)-derived
with ALLN could not be used because of the toxicity
promoter, which drives p21 expression in HT1080 p21-9
of this compound). The addition of ALLN did not
cells (Chang et al., 1999). p21 induction in GSE56-
interfere with Rb dephosphorylation, but reduced the
expressing cells resulted in complete dephosphorylation
depletion of the protein (Figure 1b), suggesting that Rb
and substantial depletion of Rb (Figure 1a, right lanes).
depletion was due to proteasome-mediated degradation.
The extent of Rb depletion was somewhat lower in the
To determine if p53 contributes to Rb depletion in
presence than in the absence of GSE56 (Figure 1a); this
response to p21, we have analysed the effects of p21 on
modest effect can be accounted for by the lower p21
Rb in HT1080 p21-9 cells, where p53 activity was
levels. Hence, p53 is not required for p21-induced Rb
blocked by GSE56, a transdominant inhibitor of p53
function (Ossovskaya et al., 1996). p21 induction by
To determine if Rb depletion is a general consequence
IPTG was about twofold lower in the presence of
of its dephosphorylation, we compared the effects of p21
GSE56 (Figure 1a, right lanes). This inhibition most
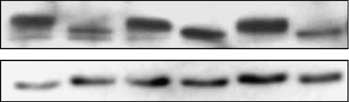
and two other Rb-dephosphorylating CDK inhibitors,p16Ink4A (CDKN1A), which inhibits CDK4/6, andp27Kip1 (CDKN2B), which inhibits CDK2. p16 andp27 were expressed in HT1080 cells from an IPTG-
IPTG (hr)
inducible retroviral vector LNXRO2 (Chang and
Roninson, 1996). IPTG-inducible expression of eitherp21 or p27 led to Rb dephosphorylation and depletion(Figure 1c). In contrast, p16 caused only dephosphory-
lation but not depletion of Rb. Hence, Rb depletion is
not a general consequence of its dephosphorylation.
We have investigated whether Rb depletion in
response to DNA damage is determined by p21.
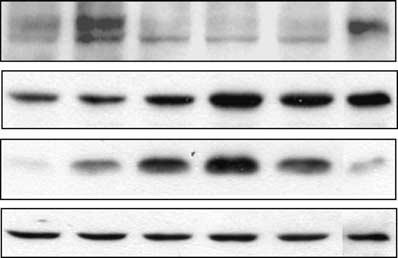
Doxorubicin treatment of HT1080 p21-9 cells induced
Mouse Rabbit
endogenous p21 and led to Rb dephosphorylation and
α-tubulin
depletion (Figure 2a, left). We have also analysed thiseffect in HT1080 p21-9 cells where p53 activity was
p21-9 GSE56
inhibited by GSE56. As shown in Figure 2a (right),GSE56 blocked p21 induction by doxorubicin almost
completely, and under these conditions, Rb showed onlyminimal dephosphorylation and no depletion. To verifythat this effect of GSE56 was directly attributable to
IPTG+ALLN
p21, we developed a derivative of HT1080 cells with
(a) Immunoblotting of p21 and Rb in HT1080 p21-9
cells with IPTG-inducible p21 (Chang et al., 1999) (left lanes) orHT1080 p21-9 transduced with p53 inhibiting peptide GSE56 (rightlanes) after the addition of 50 mM IPTG (Sigma, St Louis, MO,
USA). GSE56 (Ossovskaya et al., 1996) was introduced intoHT1080 p21-9 cells via a retroviral vector LXSE (Kandel et al.,
1997); the transduced cells were selected by flow sorting for greenfluorescent protein fluorescence. Cells were grown in Dulbecco’s
modified Eagle’s medium (DMEM) with 10% FC2 serum.
Immunoblotting was carried out by standard procedures, using
p21 mouse monoclonal antibody (Oncogene Research, San Diego,
α-tubulin
CA, USA) (top row), Rb rabbit polyclonal antibody (Neomarkers,Flemont, CA, USA) (middle row) and Rb mouse monoclonalantibody (BD PharMingen, San Diego, CA, USA) (bottom row).
a-Tubulin detected with a monoclonal antibody from Sigma wasused as a loading control. Immunoblots were developed with
horseradish peroxidase-conjugated secondary antibodies (Amer-
sham Biosciences, Piscataway, NJ, USA) and ECL chemilumines-
cence agent (Pierce, Rockford, IL, USA). Phosphorylated ( þ P)
and dephosphorylated (ÀP) forms of Rb are marked with arrows. (b) Immunoblotting of Rb in HT1080 p21-9 cells treated for 20 h
with IPTG alone or with IPTG and 20 mM proteasome inhibitorALLN (Calbiochem/EMD Biosciences, San Diego, CA, USA)
added for the last 4 h of the 20-h period is shown. Ponceau stainingwas used as a loading control. (c) Immunoblotting of Rb in
relative protein amount
HT1080 cells with IPTG-inducible expression of p21, p16 or p27,before (0 h) or after 24-h IPTG treatment (Chang and Roninson,
1996) is shown. The ratio of Rb to a-tubulin signals, as measured
by densitometry using Bio-Rad Versa Doc, is shown at the bottom. Oncogene
p21 regulates Rb degradationEV Broude et al
p21-9 GSE56 HCTp21-/- HCTp53-/-
α-tubulin Doxorubicin p21 shRNA
β-actin Doxorubicin (days) Doxorubicin
α-tubulin Doxorubicin
(a) Immunoblotting of Rb and p21 in HT1080 p21-9 cells
without (left lanes) or with GSE56 (right lanes), untreated or
treated for 16 h with 250 nM doxorubicin (Sigma). a-Tubulin wasused as a loading control. (b) Immunoblotting of Rb and p21 inHT1080-p21shRNA cells with doxycycline-inducible p21-targetingshRNA is shown. Cells grown in the absence (left lanes) or in the
presence of 100 ng/ml doxycyclin (right lanes) were treated withdoxorubicin and analysed as in (a). To derive HT1080-p21shRNA,HT1080 (from American type culture collection (ATCC) Manas-sas, VA, USA) was transduced with lentiviral vector pLV-tTR-
β-actin
KRAB-red (Wiznerowicz and Trono, 2003) (a gift from Dr DTrono; University of Geneva) expressing tTR-KRAB repressor;
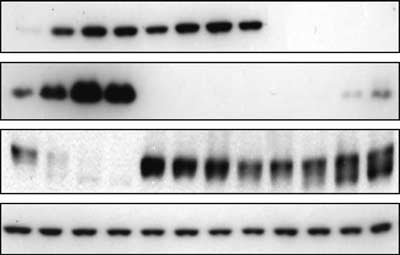
(a) Immunoblotting of p53, p21 and Rb in wild-type
infectants were selected for dsRed fluorescence. Cells were then
HCT116 cells (p53 parental) (left lanes) and HCT116 derivatives
transduced with a lentivirus expressing shRNA targeting the
with homozygous knockout of p21 (clone 80S4) (Waldman et al.,
GACCTGTCACTGTCTTGTA sequence of p21 and cloned into
1996) (middle lanes) or p53 (clone 379.2) (Bunz et al., 1998) (right
tTR-KRAB-regulated vector LLCEP TU6X (Maliyekkel et al.,
lanes), on the indicated days of treatment with 200 nM doxorubicin.
2006), followed by puromycin selection.
b-Actin (monoclonal antibody, Sigma) was used as a control. (b)Immunoblotting of Rb, p53 and p21 in HCT116 cells treated for 2
tetracycline/doxycycline-regulated expression of short
days with the indicated concentrations of doxorubicin or 2.2 nM
hairpin RNA (shRNA) against p21. shRNA induction
aphidicolin, or 2 days after exposure to ionizing radiation (6 Gy).
by doxycycline in this cell line (designated HT1080-p21shRNA) partially decreased p21 induction by
2 days of drug treatment, with only a partial decrease
doxorubicin (Figure 2b). The remaining levels of p21
on day 3 (Figure 3a). The p53À/À cell line showed no
were still sufficient to produce Rb dephosphorylation,
detectable p21 without drug treatment and only weak
but the cellular level of dephosphorylated Rb was much
p21 induction by doxorubicin (Figure 3a). In p53À/À
higher than that without p21 knockdown (Figure 2b).
cells, doxorubicin caused substantial dephosphorylation
These observations indicate that p21 mediates both
of Rb, but Rb levels were essentially unchanged
dephosphorylation and depletion of Rb in response to
(Figure 3a). This suggests that low levels of p21 induced
by the drug in p53À/À cells were sufficient to produce
To determine whether p21-mediated Rb depletion
partial dephosphorylation of Rb but not Rb depletion.
occurs in a different cell type, we investigated the effects
We next compared doxorubicin with other DNA-
of doxorubicin on Rb in HCT116 colon carcinoma cells
interacting agents that differ in their ability to induce
and in HCT116 derivatives with a homozygous knock-
p21. Doxorubicin (50 and 200 nM) and ionizing radia-
out of either p21 or p53 (Waldman et al., 1996; Bunz
tion (6 Gy) induced both p21 expression and Rb
et al., 1998). Doxorubicin treatment of parental
depletion, whereas 25 nM doxorubicin and the DNA
HCT116 cells led to both dephosphorylation and
replication inhibitor aphidicolin (2.2 nM) produced little
depletion of Rb (Figure 3a). The p21À/À line, which
or no p21 induction and did not deplete Rb (Figure 3b).
expresses p53 but not p21, expressed much higher basal
These results confirm the role of p21 in inducing Rb
levels of Rb than the wild-type cells, and it showed
dephosphorylation and degradation in response to
no Rb dephosphorylation and no decrease in Rb after
Oncogene
In summary, we have found that p21 (which inhibits
The sequence of events, wherein p21 first activates Rb
CDK2 and CDK4/6) and p27 (a selective CDK2
through dephosporylation and then deactivates it
inhibitor) cause not only dephosphorylation but also
through depletion, suggests negative feedback regula-
degradation of Rb. The finding that p16 (a selective
tion that limits the duration of Rb-mediated checkpoint
CDK4/6 inhibitor) does not deplete Rb would at first
arrest. It is noteworthy that senescent (permanently
glance suggest that phosphorylation by CDK2, but not
arrested) fibroblasts express primarily p16 rather than
by CDK4/6, protects Rb from proteolytic degradation.
p21, and p16 does not cause Rb degradation. We note,
However, this interpretation seems unlikely, because
however, that this pattern is not common to all
cellular expression of p16 was shown to inhibit not only
senescent fibroblast cultures, since some senescent
CDK4/6 but also CDK2, by causing reassortment of
fibroblasts show apparent Rb depletion by immuno-
cyclin–CDK-inhibitor complexes (Jiang et al., 1998). On
blotting (Stein et al., 1999). In contrast to normal
the other hand, p21 and p27 are structurally much
senescent fibroblasts, p21 expression is maintained over
more similar to each other than to p16 outside their
a long term in p16-deficient tumor cells rendered
CDK-binding domains, and it is plausible that their
senescent by DNA damage (Chang et al., 2002), and
interactions with non-CDK proteins are responsible
in such cases Rb appears dispensable for long-term
for the degradation of dephosphorylated Rb. This
maintenance of cell-cycle arrest. p21-mediated transcrip-
interpretation is also consistent with the observed
tional inhibition of multiple cell-cycle progression genes
dephosphorylation but not degradation of Rb in cells
(Chang et al., 2000) provides one of the mechanisms that
expressing low levels of p21 (Figures 2a and 3a). The
can sustain cell-cycle arrest in the absence of Rb, and
requirement for additional events beyond dephosphory-
additional mechanisms will undoubtedly be elucidated
lation can explain why the depletion of dephosphorylated
Rb is detectable in some, but not all, human cell lines
p21 is almost never mutated in cancers, and its pattern
exposed to DNA damage (see, for example, Figure 2
of expression in human tumors is consistent with a role
of both a tumor suppressor and an oncogene (Roninson,
Rb degradation is likely to be mediated by the
2002). The finding that p21 both activates Rb by
proteasome, since it is inhibited by ALLN that has
dephosporylation and inactivates it by degradation
proteasome inhibition as its primary effect (Drexler,
provides another explanation for the dual tumor
1997). p21-induced Rb degradation is unlikely to be
suppressor/oncogenic functions of p21.
mediated by Mdm2, because it was not prevented by theinhibition of p53, a positive regulator of Mdm2.
Although there is no evidence for a direct bindingbetween p21 and Rb, p21 was reported to be associated
We thank Dr Bert Vogelstein for HCT116 derivatives, Dr
with 20S proteasome that also degrades Rb (Sdek et al.,
Didier Trono for pLV-tTR-KRAB-red and Natalie Warholic
2005), and it is conceivable that p21 could drive some as
and Jennifer Huntington for technical assistance. This work
yet unidentified Rb-containing multimeric complexes to
was supported by NIH grants R01 AG17921, R01 CA62099
Boyer SN, Wazer DE, Band V. (1996). E7 protein of human
human tumor cells by chemotherapeutic drugs. Oncogene
papilloma virus-16 induces degradation of retinoblastoma
protein through the ubiquitin–proteasome pathway. Cancer
Drexler HC. (1997). Activation of the cell death program by
inhibition of proteasome function. Proc Natl Acad Sci USA
Bunz F, Dutriaux A, Lengauer C, Waldman T, Zhou S, Brown
JP et al. (1998). Requirement for p53 and p21 to sustain G2
Higashitsuji H, Itoh K, Nagao T, Dawson S, Nonoguchi K,
arrest after DNA damage. Science 282: 1497–1501.
Kido T et al. (2000). Reduced stability of retinoblastoma
Chang BD, Roninson IB. (1996). Inducible retroviral vectors
protein by gankyrin, an oncogenic ankyrin-repeat protein
regulated by lac repressor in mammalian cells. Gene 183:
overexpressed in hepatomas. Nat Med 6: 96–99.
Jiang H, Chou HS, Zhu L. (1998). Requirement of cyclin
Chang BD, Swift ME, Shen M, Fang J, Broude EV, Roninson
E-Cdk2 inhibition in p16(INK4a)-mediated growth sup-
IB. (2002). Molecular determinants of terminal growth
pression. Mol Cell Biol 18: 5284–5290.
arrest induced in tumor cells by a chemotherapeutic drug.
Kandel ES, Chang BD, Schott B, Shtil AA, Gudkov AV,
Proc Natl Acad Sci USA 99: 389–394.
Roninson IB. (1997). Applications of green fluorescent
Chang BD, Watanabe K, Broude EV, Fang J, Poole JC,
protein as a marker of retroviral vectors. Somat Cell Mol
Kalinichenko TV et al. (2000). Effects of p21Waf1/Cip1/Sdi1 on
cellular gene expression: implications for carcinogenesis,
Kehn K, Fuente CL, Strouss K, Berro R, Jiang H, Brady J
senescence, and age-related diseases. Proc Natl Acad Sci
et al. (2005). The HTLV-ITax oncoprotein targets the
retinoblastoma protein for proteasomal degradation. Onco-
Chang BD, Xuan Y, Broude EV, Zhu H, Schott B,
Fang J et al. (1999). Role of p53 and p21waf1/cip1 in
Maliyekkel A, Davis BA, Roninson IB. (2006). Cell cycle
senescence-like terminal proliferation arrest induced in
arrest drastically extends the duration of gene silencing after
Oncogene
p21 regulates Rb degradationEV Broude et al
transient expression of short hairpin RNA. Cell Cycle 5:
Stein GH, Drullinger LF, Soulard A, Dulic V. (1999).
Differential roles for cyclin-dependent kinase inhibitors
Ossovskaya VS, Mazo IA, Chernov MV, Chernova OB, Strezoska
p21 and p16 in the mechanisms of senescence and
Z, Kondratov R et al. (1996). Use of genetic suppressor
differentiation in human fibroblasts. Mol Cell Biol 19:
elements to dissect distinct biological effects of separate p53
domains. Proc Natl Acad Sci USA 93: 10309–10314.
Tan X, Martin SJ, Green DR, Wang JY. (1997). Degradation
Roninson IB. (2002). Oncogenic functions of tumour suppres-
retinoblastoma protein in tumor necrosis factor-
sor p21(Waf1/Cip1/Sdi1): association with cell senescence
and tumour-promoting activities of stromal fibroblasts.
Waldman T, Lengauer C, Kinzler KW, Vogelstein B. (1996).
Sdek P, Ying H, Chang DL, Qiu W, Zheng H, Touitou R et al.
Uncoupling of S phase and mitosis induced by anticancer
(2005). MDM2 promotes proteasome-dependent ubiquitin-
agents in cells lacking p21. Nature 381: 713–716.
independent degradation of retinoblastoma protein. Mol
Wiznerowicz M, Trono D. (2003). Conditional suppression of
cellular genes: lentivirus vector-mediated drug-inducible
Smith ML, Zhan Q, Bae I, Fornace Jr AJ. (1994). Role of
RNA interference. J Virol 77: 8957–8961.
retinoblastoma gene product in p53-mediated DNA damage
Ying H, Xiao ZX. (2006). Targeting retinoblastoma protein
response. Exp Cell Res 215: 386–389.
for degradation by proteasomes. Cell Cycle 5: 506–508. Oncogene
CODIGO DE ETICA DEL PROFESOR AMBITO DE APLICACION, OBJETO, PRINCIPIOS FUNDAMENTALES CAPÍTULO I ÁMBITO DE APLICACIÓN Artículo 1. El presente instrumento jurídico contiene el Código de Ética Docente de la Facultad de Derecho y Ciencias Políticas de la Universidad Nacional de Pilar, que CAPÍTULO II Artículo 2. El objeto de este Código es establecer un conjunto
Informationen zur Neuen Grippe (Influenza A/H1N1) für Schulleitungen 1. Situationseinschätzung Deutschland Das Virus A/H1N1, das die Neue Grippe verursacht, kann leicht von Mensch zu Mensch übertragen werden. Das zeigt die ständig steigende Zahl der Erkrankten. Derzeit sind in Deutschland 16.835 Personen an der Neu-en Grippe erkrankt (Stand 04. September 2009). Die Mehrzahl d
 Oncogene (2007), 1–5& 2007 Nature Publishing Group All rights reserved 0950-9232/07 $30.00
p21Waf1/Cip1/Sdi1 mediates retinoblastoma protein degradation
EV Broude1, ME Swift2, C Vivo1, B-D Chang1, BM Davis1, S Kalurupalle1, MV Blagosklonny1and IB Roninson1
1Cancer Center, Ordway Research Institute, Albany, NY, USA and 2Department of Molecular Genetics, University of Illinois atChicago, Chicago, IL, USA
Damage-induced G1 checkpoint in mammalian cells
the proteasome. Proteasome-mediated Rb degradation
involves upregulation of p53, which activates transcription
is promoted by Mdm2 (Sdek et al., 2005) and gankyrin
of p21Waf1 (CDKN1A). Inhibition of cyclin-dependent
(Higashitsuji et al., 2000), E7 of papilloma virus (Boyer
kinase (CDK)2 and CDK4/6 by p21 leads to depho-
et al., 1996) and Tax of HTLV1 (Kehn et al., 2005).
Oncogene (2007), 1–5& 2007 Nature Publishing Group All rights reserved 0950-9232/07 $30.00
p21Waf1/Cip1/Sdi1 mediates retinoblastoma protein degradation
EV Broude1, ME Swift2, C Vivo1, B-D Chang1, BM Davis1, S Kalurupalle1, MV Blagosklonny1and IB Roninson1
1Cancer Center, Ordway Research Institute, Albany, NY, USA and 2Department of Molecular Genetics, University of Illinois atChicago, Chicago, IL, USA
Damage-induced G1 checkpoint in mammalian cells
the proteasome. Proteasome-mediated Rb degradation
involves upregulation of p53, which activates transcription
is promoted by Mdm2 (Sdek et al., 2005) and gankyrin
of p21Waf1 (CDKN1A). Inhibition of cyclin-dependent
(Higashitsuji et al., 2000), E7 of papilloma virus (Boyer
kinase (CDK)2 and CDK4/6 by p21 leads to depho-
et al., 1996) and Tax of HTLV1 (Kehn et al., 2005).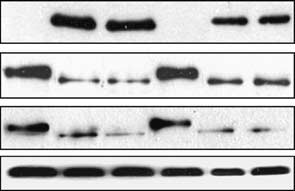




 treated with IPTG for 20 h with ALLN added in one
likely reflects the effect of GSE56 on the p53 response
arm of the experiment for the last 4 h (longer treatment
element present in the cytomegalovirus (CMV)-derived
with ALLN could not be used because of the toxicity
promoter, which drives p21 expression in HT1080 p21-9
of this compound). The addition of ALLN did not
cells (Chang et al., 1999). p21 induction in GSE56-
interfere with Rb dephosphorylation, but reduced the
expressing cells resulted in complete dephosphorylation
depletion of the protein (Figure 1b), suggesting that Rb
and substantial depletion of Rb (Figure 1a, right lanes).
treated with IPTG for 20 h with ALLN added in one
likely reflects the effect of GSE56 on the p53 response
arm of the experiment for the last 4 h (longer treatment
element present in the cytomegalovirus (CMV)-derived
with ALLN could not be used because of the toxicity
promoter, which drives p21 expression in HT1080 p21-9
of this compound). The addition of ALLN did not
cells (Chang et al., 1999). p21 induction in GSE56-
interfere with Rb dephosphorylation, but reduced the
expressing cells resulted in complete dephosphorylation
depletion of the protein (Figure 1b), suggesting that Rb
and substantial depletion of Rb (Figure 1a, right lanes).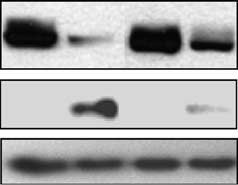
 p21 regulates Rb degradationEV Broude et al
p21-9 GSE56
p21 regulates Rb degradationEV Broude et al
p21-9 GSE56 In summary, we have found that p21 (which inhibits
The sequence of events, wherein p21 first activates Rb
CDK2 and CDK4/6) and p27 (a selective CDK2
through dephosporylation and then deactivates it
inhibitor) cause not only dephosphorylation but also
through depletion, suggests negative feedback regula-
degradation of Rb. The finding that p16 (a selective
tion that limits the duration of Rb-mediated checkpoint
CDK4/6 inhibitor) does not deplete Rb would at first
arrest. It is noteworthy that senescent (permanently
glance suggest that phosphorylation by CDK2, but not
arrested) fibroblasts express primarily p16 rather than
by CDK4/6, protects Rb from proteolytic degradation.
In summary, we have found that p21 (which inhibits
The sequence of events, wherein p21 first activates Rb
CDK2 and CDK4/6) and p27 (a selective CDK2
through dephosporylation and then deactivates it
inhibitor) cause not only dephosphorylation but also
through depletion, suggests negative feedback regula-
degradation of Rb. The finding that p16 (a selective
tion that limits the duration of Rb-mediated checkpoint
CDK4/6 inhibitor) does not deplete Rb would at first
arrest. It is noteworthy that senescent (permanently
glance suggest that phosphorylation by CDK2, but not
arrested) fibroblasts express primarily p16 rather than
by CDK4/6, protects Rb from proteolytic degradation. p21 regulates Rb degradationEV Broude et al
transient expression of short hairpin RNA. Cell Cycle 5:
Stein GH, Drullinger LF, Soulard A, Dulic V. (1999).
p21 regulates Rb degradationEV Broude et al
transient expression of short hairpin RNA. Cell Cycle 5:
Stein GH, Drullinger LF, Soulard A, Dulic V. (1999).