Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Pntd.0000076 1.3
Ivermectin Resistance in Onchocerca volvulus: Towarda Genetic Basis
1 Laboratory of Molecular Parasitology, Lindsley F. Kimball Research Institute, New York Blood Center, New York, New York, United States of America, 2 Divergence Inc., St.
Louis, Missouri, United States of America, 3 Department of Genetics, Washington University School of Medicine, St. Louis, Missouri, United States of America
parasite transmission, possibly with IVM-resistant O. volvulusleading to disease recrudescence. Additionally, studies have
Onchocerciasis (river blindness) is a human disease caused by
the filarial worm Onchocerca volvulus. Adult worms can live for over
particularly the b-tubulin gene in human O. volvulus and the
a decade in skin nodules of affected humans, releasing millions of
livestock nematode parasite Haemonchus contortus [22,26]. However,
microfilariae that cause debilitating itching and blindness [1]. An
previous O. volvulus genotyping studies were non-longitudinal,
estimated 37 million people are infected [2], and there are 46,000
using worms collected from different IVM-naı¨ve and treated
new cases of blindness annually (http://www.apoc.bf/).
International programs supported by the World Health
Organization and many other groups have worked to control
A New Study: IVM Causes Genetic Selection on
the impact of onchocerciasis using vector control with insecticides
beginning in 1974 and mass drug administration (MDA) withivermectin (IVM, brand name Mectizan) beginning in 1987
A new study by Catherine Bourguinat and colleagues published
(Figure 1) [3]. IVM is a highly effective microfilaricide and inhibits
in PLoS Neglected Tropical Diseases extends these previous reports and
female worm microfilarial production for several months. Annual
concludes not only that IVM causes genetic selection on O. volvulus
IVM MDA reduces morbidity [4,5] and lowers transmission [6,7].
worms, but that this selection is also associated with a lower
From 1974 to 2002, the Onchocerciasis Control Programme
reproductive rate of the female parasites [27]. In this study of O.
(OCP) in West Africa greatly decreased O. volvulus transmission in
volvulus treatment in a hyperendemic region of central Cameroon,
the 11 OCP countries and prevented 600,000 cases of blindness
parasite genotypes (b-tubulin gene and two controls) and pheno-
[8–10]. IVM without vector control has been the principal tool for
types (female fertility) were characterized in worms collected from
the Onchocerciasis Elimination Program of the Americas (1992–
the same individuals before and after four or 13 IVM treatments
present) [9] and the African Programme for Onchocerciasis
over three years. Parasites were collected pre- and post-treatment
Control (1995–present). In the Americas, where O. volvulus is less
from clinical trial patients in four IVM treatment groups: 150 mg/
common, the Onchocerciasis Elimination Program has substan-
kg of body weight annually or three-monthly, and 800 mg/kg
tially reduced transmission and is on track to eliminate the disease
Analyses of the genetic polymorphism in parasites pre- and post-
The African Programme for Onchocerciasis Control has
treatment clearly showed a significant selection for b-tubulin
extended treatment to 19 countries beyond those originally
heterozygotes in female worms. The most marked effect was in
included in the OCP through sustainable community-directed
the three-monthly treated groups, where the frequency of the b-
IVM treatment [1,11]. By the end of 2005, 400 million treatments
tubulin ‘‘aa’’ homozygotes post-IVM was reduced on average from
had been supplied in Africa by Merck’s Mectizan Donation
68.6% to 25.6%, while the ‘‘ab’’ heterozygotes increased from
Program, with an estimated 40 million people treated by nearly
20.9% to 69.2% over three years. Moreover, b-tubulin ‘‘aa’’
300,000 community distributors (http://www.apoc.bf/). Never-
homozygous females were significantly more fertile than hetero-
theless, the ecology of the disease in Africa, including the broad
zygotes before treatment (67% versus 37%) and 12 months
geographic range of O. volvulus and its blackfly vector, leads to theestimation that IVM treatment of at least 65% of the population
after the last IVM dose in the groups treated annually (60%
for 25 or more years will be necessary to eliminate infection [9,12].
versus 17%). No significant selection was observed in the control
There are significant logistical obstacles to achieving such
broad-ranging and prolonged treatment, and there is also concernthat O. volvulus resistance to IVM will emerge. IVM resistancehas become widespread in many parasitic nematodes of
Citation: Lustigman S, McCarter JP (2007) Ivermectin Resistance in Onchocerca
livestock [13,14]. At present there are no alternative drugs for
volvulus: Toward a Genetic Basis. PLoS Negl Trop Dis 1(1): e76. doi:10.1371/journal. pntd.0000076
IVM for use in the Onchocerca MDA programs that reducemicrofilariae or kill adult worms, which can live up to 15 years in
Received July 17, 2007; Accepted August 1, 2007; Published August 30, 2007
Copyright: ß 2007 Lustigman, McCarter. This is an open-access article
The emergence of drug-resistant O. volvulus has been suggested
distributed under the terms of the Creative Commons Attribution License, whichpermits unrestricted use, distribution, and reproduction in any medium, provided
by reports of patients failing to respond to IVM treatment [15,16].
the original author and source are credited.
A recent report from Ghana has provided the first proof of IVM
Funding: The authors received no specific funding for this study.
resistance in O. volvulus: Mike Osei-Atweneboana and colleagues
Competing Interests: SL has received research grants for the development of
showed that the ability of IVM to suppress skin microfilariae
anti-Onchocerca vaccines and macrofilaricides. JPM is a share-holder and
repopulation was reduced in some communities that had received
employee of Divergence, Inc., a company that works on parasite control in
6–18 years of IVM MDA [17]. The authors predict that a high
rate of repopulation of skin with microfilariae will allow continued
PLoS Neglected Tropical Diseases | www.plosntds.org
Figure 1. Distribution of Onchocerciasis Showing Current Status of Global Onchocerciasis Control. Red shading represents areasreceiving ivermectin treatment. Yellow shading represents areas requiring further epidemiological surveys. Green shading indicates the area coveredby the OCP in West Africa. Pink zones indicate the special intervention zones, i.e., previous OCP areas receiving ivermectin and some vector control. Figure from [10]. doi:10.1371/journal.pntd.0000076.g001
important to examine for polymorphisms in these genes in theuniquely collected O. volvulus female worms described in this study
A major strength of this study is that the O. volvulus parasites
were collected from the same individuals before and after IVM
Despite these caveats, the study indicates that IVM causes
treatments. Therefore the observed changes in genotype frequen-
genetic selection on O. volvulus worms and points to the daunting
cies between IVM-naı¨ve and treated O. volvulus populations are not
possibility of the spread of IVM-resistant parasites in endemic
due to factors such as geographical or sampling effects.
regions that have been treated with IVM.
The main limitation of this study was that some worm samples
could not be genotyped, thus reducing the number that could be
Implications of the Study for River Blindness
analyzed, particularly after treatment. This limitation might also
have impeded the genotyping of DNA, ideally prepared fromworm sections instead of just whole females. What is given as
The finding that IVM treatment selected for b-tubulin hetero-
a single genotype is, in fact, a consensus of multiple genotypes
zygotes and that this selection was dependent on dosage raises
including the adult female body and progeny (uterine embryos and
important concerns for the current river blindness control
microfilariae). Furthermore, the samples from unfertile females,
programs. These concerns are heightened by the fact that this
which probably represent true singletons, were treated the same as
gene has been linked with IVM resistance in another parasitic
those from females classified as being of low or high fertility.
nematode [26], and by the recent evidence that IVM resistance is
Consequently, the study leaves unanswered questions including
occurring in O. volvulus [17]. Semiannual or more frequent
whether the selection of the b-tubulin heterozygote genotypes is on
treatments are ongoing in some endemic areas and are under
the females or their progeny. If it is on the progeny, questions
consideration in other areas. Such treatment might increase the
remain regarding the fitness and susceptibility to IVM treatment
selection pressure. Therefore, Bourguinat and colleagues’ study is
of the b-tubulin heterozygote microfilariae once they develop into
a wake-up call for control programs to select their treatment
the infective stage larvae and enter a new human host.
regimens carefully and to develop plans for detecting IVM
The authors do not present a hypothesis to explain why IVM
resistance and the associated genetic markers (control programs
causes selection for b-tubulin heterozygote genotypes. The
will require additional funding for these plans). This study presents
glutamate-gated chloride channels are thought to be involved in
a possible structure of study design that will incorporate the
the mode of action of IVM and resistance to the drug [28].
detection and validation of the genetic markers associated with
Treatment with IVM is known to cause a loss of polymorphism
not only at certain b-tubulin gene loci, but also at certain loci of the
Simultaneously, we need to greatly increase our current level of
genes encoding the gamma-aminobutyric acid receptor, gluta-
effort and support to develop and test a new generation of control
mate-gated chloride channel, and ATP-binding cassette trans-
tools for onchocerciasis. These tools should include both vaccines
porter of IVM-resistant H. contortus [23]. It will therefore be
and macrofilaricides (drugs which kill adult worms) that have new
PLoS Neglected Tropical Diseases | www.plosntds.org
classes of chemistry with novel modes of action. Recent break-
McConnell Clark Foundation [31]. It may be possible to link such
throughs now make macrofilaricide development more feasible,
a vaccine with drug treatments in a program of vaccine-linked
and accordingly such development is now a high-priority goal with
chemotherapy [32,33]. These new generations of control tools
the World Health Organization’s Special Programme for Re-
would complement the present control measure—the establish-
search and Training in Tropical Diseases and the Bill and Melinda
ment of sustainable community-directed treatment with IVM—
Gates Foundation [29,30]. The development of an anti-Onchocerca
and ultimately support the long-term goal of eliminating
vaccine has been the focus of research supported by the Edna
onchocerciasis as a public health problem in Sub-Saharan Africa.
1. Richards FO Jr, Boatin B, Sauerbrey M, Seketeli A (2001) Control of
in endemic communities in Ghana: A two-phase epidemiological study. Lancet
onchocerciasis today: status and challenges. Trends Parasitol 17: 558–563.
2. African Programme for Onchocerciasis Control (2005) Final communique´ of the
18. Kohler P (2001) The biochemical basis of anthelmintic action and resistance.
11th session of the Joint Action Forum (JAF) of APOC, Paris, France, 6–9
December 2005. Available: http://www.apoc.bf/en/download.htm. Accessed 3
19. Huang YJ, Prichard RK (1999) Identification and stage-specific expression of
two putative P-glycoprotein coding genes in Onchocerca volvulus. Mol Biochem
3. Peters DH, Phillips T (2004) Mectizan Donation Program: Evaluation of
a public–private partnership. Trop Med Int Health 9: A4–A15.
20. Ardelli BF, Guerriero SB, Prichard RK (2005) Genomic organization and effects
4. Ejere H, Schwartz E, Wormald R (2001) Ivermectin for onchocercal eye disease
of ivermectin selection on Onchocerca volvulus P-glycoprotein. Mol Biochem
(river blindness). Cochrane Database Syst Rev: CD002219.
5. Tielsch JM, Beeche A (2004) Impact of ivermectin on illness and disability
21. Bourguinat C, Pion SD, Kamgno J, Gardon J, Gardon-Wendel N, et al. (2006)
associated with onchocerciasis. Trop Med Int Health 9: A45–A56.
Genetic polymorphism of the beta-tubulin gene of Onchocerca volvulus in
6. Boussinesq M, Prod’hon J, Chippaux JP (1997) Onchocerca volvulus: Striking
ivermectin naive patients from Cameroon, and its relationship with fertility of
decrease in transmission in the Vina valley (Cameroon) after eight annual large
scale ivermectin treatments. Trans R Soc Trop Med Hyg 91: 82–86.
22. Eng JK, Prichard RK (2005) A comparison of genetic polymorphism in
7. Collins RC, Gonzales-Peralta C, Castro J, Zea-Flores G, Cupp MS, et al. (1992)
populations of Onchocerca volvulus from untreated- and ivermectin-treated patients.
Ivermectin: Reduction in prevalence and infection intensity of Onchocerca volvulus
Mol Biochem Parasitol 142: 193–202.
following biannual treatments in five Guatemalan communities. Am J Trop Med
23. Ardelli BF, Prichard RK (2004) Identification of variant ABC-transporter genes
among Onchocerca volvulus collected from ivermectin-treated and untreated
8. Molyneux DH (1995) Onchocerciasis control in West Africa: Current status and
patients in Ghana, West Africa. Ann Trop Med Parasitol 98: 371–384.
future of the Onchocerciasis Control Programme. Parasitol Today 11: 399–402.
24. Ardelli BF, Guerriero SB, Prichard RK (2006) Ivermectin imposes selection
9. Boatin BA, Richards FO Jr (2006) Control of onchocerciasis. Adv Parasitol 61:
pressure on P-glycoprotein from Onchocerca volvulus: Linkage disequilibrium and
genotype diversity. Parasitology 132: 375–386.
10. Basa´n˜ez MG, Pion SD, Churcher TS, Breitling LP, Little MP, et al. (2006) River
25. Ardelli BF, Guerriero SB, Prichard RK (2006) Characterization of a half-size
blindness: A success story under threat? PLoS Med 3: e371. doi:10.1371/
ATP-binding cassette transporter gene which may be a useful marker for
ivermectin selection in Onchocerca volvulus. Mol Biochem Parasitol 145: 94–100.
11. Remme JHF (1995) The African Programme for Onchocerciasis Control:
26. Eng JK, Blackhall WJ, Osei-Atweneboana MY, Bourguinat C, Galazzo D, et al.
Preparing to launch. Parasitol Today 11: 403–406.
(2006) Ivermectin selection on beta-tubulin: Evidence in Onchocerca volvulus and
12. Winnen M, Plaisier AP, Alley ES, Nagelkerke NJ, van Oortmarssen G, et al.
Haemonchus contortus. Mol Biochem Parasitol 150: 229–235.
(2002) Can ivermectin mass treatments eliminate onchocerciasis in Africa? BullWorld Health Organ 80: 384–391.
27. Bourguinat C, Pion SDS, Kamgno J, Gardon J, Duke BOL, et al. (2007) Genetic
13. Coles GC (2006) Drug resistance and drug tolerance in parasites. Trends
selection of low fertile Onchocerca volvulus by ivermectin treatment. PLoS Negl
Parasitol 22: 348; author reply 349.
Trop Dis 1: e72. doi:10.1371/journal.pntd.0000072
14. Coles GC, Jackson F, Pomroy WE, Prichard RK, von Samson-Himmelstjerna G,
28. Njue AI, Prichard RK (2004) Genetic variability of glutamate-gated chloride
et al. (2006) The detection of anthelmintic resistance in nematodes of veterinary
channel genes in ivermectin-susceptible and -resistant strains of Cooperia oncophora.
importance. Vet Parasitol 136: 167–185.
15. Awadzi K, Attah SK, Addy ET, Opoku NO, Quartey BT, et al. (2004) Thirty-
29. Behm CA, Bendig MM, McCarter JP, Sluder AE (2005) RNAi-based discovery
month follow-up of sub-optimal responders to multiple treatments with
and validation of new drug targets in filarial nematodes. Trends Parasitol 21:
ivermectin, in two onchocerciasis-endemic foci in Ghana. Ann Trop Med
30. Nwaka S, Hudson A (2006) Innovative lead discovery strategies for tropical
16. Awadzi K, Boakye DA, Edwards G, Opoku NO, Attah SK, et al. (2004) An
diseases. Nat Rev Drug Discov 5: 941–955.
investigation of persistent microfilaridermias despite multiple treatments with
31. Cook JA, Steel C, Ottesen EA (2001) Towards a vaccine for onchocerciasis.
ivermectin, in two onchocerciasis-endemic foci in Ghana. Ann Trop Med
32. Hotez PJ, Ferris MT (2006) The antipoverty vaccines. Vaccine 24: 5787–5799.
17. Osei-Atweneboana MY, Eng JK, Boakye DA, Gyapong JO, Prichard RK (2007)
33. Hotez PJ (2007) Control of onchocerciasis—The next generation. Lancet 369:
Prevalence and intensity of Onchocerca volvulus infection and efficacy of ivermectin
PLoS Neglected Tropical Diseases | www.plosntds.org
Checkliste – Einweisung Diabetes-Klinik Für Kinder-/Jugend-Kurse, Pumpen-Kurse bitte extra Checkliste und zusätzliche Anmeldeunterlagen verwenden, für Spezial-Gruppen bitte diese Checkliste und zusätzliche Anmeldeunterlagen verwenden (sh. www.diabetes-zentrum.de/downloads.html oder Telefon 07931 594-101). Wahlleistungen Termin am ________________________ Pfl
PROGRAM FOR KONFERENCEN FM I STRATEGISK LEDELSEVelkomst og indledning Birgitte Dyrvig Carlsson, afdelingsleder, Cowi A/S, ordstyrer, bestyrelsesmedlem, DFM-netværk Facilities Management en strategisk ledelsesdisciplinHovedelementerne i Facilities Management og beskrivelse af, hvordan Facilities Management har Ændring fra udbygningsfokuseret FM til FM som integreret del af museets strategiske
 Ivermectin Resistance in Onchocerca volvulus: Towarda Genetic Basis
1 Laboratory of Molecular Parasitology, Lindsley F. Kimball Research Institute, New York Blood Center, New York, New York, United States of America, 2 Divergence Inc., St.
Ivermectin Resistance in Onchocerca volvulus: Towarda Genetic Basis
1 Laboratory of Molecular Parasitology, Lindsley F. Kimball Research Institute, New York Blood Center, New York, New York, United States of America, 2 Divergence Inc., St.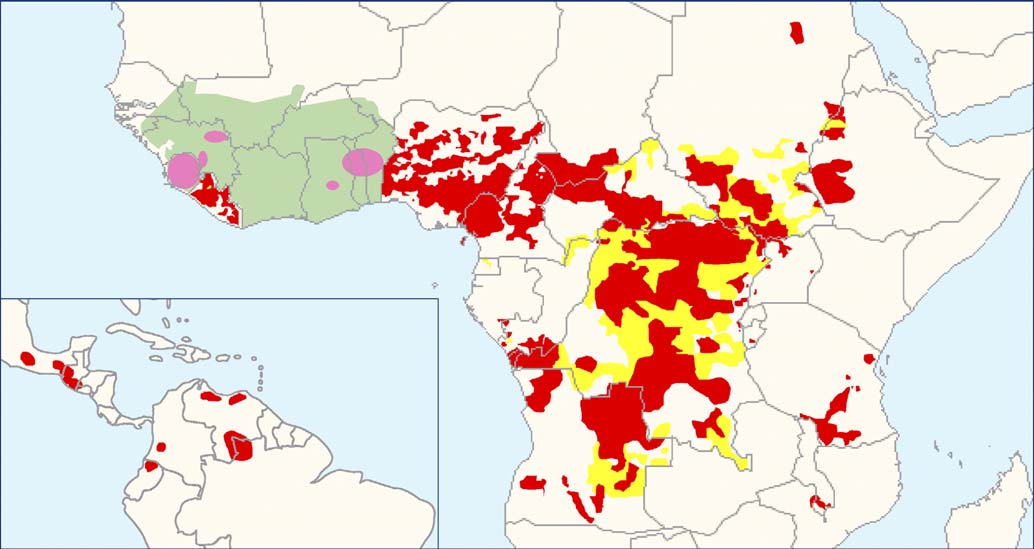
 Figure 1. Distribution of Onchocerciasis Showing Current Status of Global Onchocerciasis Control. Red shading represents areasreceiving ivermectin treatment. Yellow shading represents areas requiring further epidemiological surveys. Green shading indicates the area coveredby the OCP in West Africa. Pink zones indicate the special intervention zones, i.e., previous OCP areas receiving ivermectin and some vector control.
Figure 1. Distribution of Onchocerciasis Showing Current Status of Global Onchocerciasis Control. Red shading represents areasreceiving ivermectin treatment. Yellow shading represents areas requiring further epidemiological surveys. Green shading indicates the area coveredby the OCP in West Africa. Pink zones indicate the special intervention zones, i.e., previous OCP areas receiving ivermectin and some vector control. classes of chemistry with novel modes of action. Recent break-
McConnell Clark Foundation [31]. It may be possible to link such
throughs now make macrofilaricide development more feasible,
a vaccine with drug treatments in a program of vaccine-linked
and accordingly such development is now a high-priority goal with
chemotherapy [32,33]. These new generations of control tools
the World Health Organization’s Special Programme for Re-
would complement the present control measure—the establish-
search and Training in Tropical Diseases and the Bill and Melinda
ment of sustainable community-directed treatment with IVM—
Gates Foundation [29,30]. The development of an anti-Onchocerca
and ultimately support the long-term goal of eliminating
vaccine has been the focus of research supported by the Edna
onchocerciasis as a public health problem in Sub-Saharan Africa.
classes of chemistry with novel modes of action. Recent break-
McConnell Clark Foundation [31]. It may be possible to link such
throughs now make macrofilaricide development more feasible,
a vaccine with drug treatments in a program of vaccine-linked
and accordingly such development is now a high-priority goal with
chemotherapy [32,33]. These new generations of control tools
the World Health Organization’s Special Programme for Re-
would complement the present control measure—the establish-
search and Training in Tropical Diseases and the Bill and Melinda
ment of sustainable community-directed treatment with IVM—
Gates Foundation [29,30]. The development of an anti-Onchocerca
and ultimately support the long-term goal of eliminating
vaccine has been the focus of research supported by the Edna
onchocerciasis as a public health problem in Sub-Saharan Africa.