Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Fm t52-2012.indd
ŁUKASZ NOWAK1, DANIEL ŻUROWSKI1, JAN DOBROGOWSKI3, JERZY WORDLICZEK2,
PENTOXIFYLLINE MODIFIES CENTRAL AND PERIPHERAL
VAGAL MECHANISM IN ACUTE AND CHRONIC PAIN MODELS
Abstract: Pentoxifylline modifi es central and peripheral vagal mechanism in acute and chronic pain models.
The infl ammatory process gives the way to hyperalgesia that is documented by the animal experimental studies. Pentoxifylline (PTX) has strong antyinfl amatory effects, decreases TNF-alpha and other proinfl ammatory cytokines production. Therefore, the aim of present investigation was to evaluate the effectiveness of PTX in nociception processes, especially in aspects of vagal activity, in experimental pain models: visceral pain (VP), neuropathy (CCI) and neurogenic infl ammation (NI). In VP and CCI models we observed signifi cant increase in the pain threshold after blocking proinfl ammatory cytokines whereas in NI there was no such effect. In our studies we also observed the increase of vagal afferents activity in VP and CCI, on the contrary to NI model. In summary, our study demonstrates that preemptive inhibition of proinfl ammatory cytokine synthesis by treatment with PTX is useful in antagonizing hyperalgesia in infl ammatory pain. Pentoxifylline reduces central and peripheral sensitization processes depend on the vagal component in both acute and chronic pain models but in a different manner and mechanisms. Our results establish the participation of infl ammatory and vagal component in nociception. The modulation of the vagal system offers the new possibilities of the pain treatment in patients resistant to the classical analgesic therapy. Key words: pentoxifylline, visceral pain, neuropathy, neurogenic infl ammation, vagus nerve
There is increasing evidence showing that sensitization processes are not only controlled by spinal afferent fi bers but it is also related to vagal afferent fi bers [1–4]. Hypersensitivity in response to mechanical and chemical stimuli results from the sensitization of the primary afferent neurons (peripheral hyperalgesia)
and the nociceptive pathways in the central nervous system (CNS) (central hyperalgesia) [5, 6]. The vagus nerve, the largest sensory nerve in the body, and its involvement in infl ammatory response, can be a target for anti-pain therapy lately, due to growing evidence that indicates its contribution in nociception.
Vagotomy studies demonstrate that vagal afferent integrity is essential to
the effi cacy of different analgesic treatments like morphine [7]. Furthermore, subdiaphragmatic vagotomy decreases the threshold for paw withdrawal in rats [8], increases sensitivity to noxious lesions [9] and enhances hyperalgesia induced by the potent infl ammatory mediator bradykinin [10, 11]. However, vagotomy also prevents the establishment of kainic acid-induced hyperalgesia in mice [12] and reduces nociception in the formalin test in male rats [13]. On the other hand an anti-nociceptive action of VNS is reported in numerous studies which describe for instance inhibition of the nociceptive refl ex [14, 15] or latency increase of the tail-fl ick response to heat in rats [16]. However, pronociceptive effects have also been reported in the tail-fl ick test in rats [17]. These discrepancies can be explained by differences in stimulation parameters : low intensity stimulations of vagal afferents facilitate, while high intensity stimulations inhibit nociception [7]. According to Ren et al., the analgesic effect of VNS seems to depend on a critical stimulation intensity that activates C-fi bers [18].
Pentoxifylline (PTX), a nonspecifi c phosphodiesterase inhibitor, has several
actions that improve blood rheology and tissue perfusion and it is used clinically as a treatment for intermittent claudication [19]. Several laboratory investigations, however, employ the drug for its anticytokine effects. Previously several reports demonstrate that PTX administration virtually eliminates the up-regulation of cytokines in various pain models [20–26]. Its infl uence on infl ammatory component of sensitization processes is well known, but on the vagal dependent mechanisms, remain unclear [27]. Thus the aim of present investigation was to evaluate the effectiveness of PTX in peripheral and central sensitization processes, especially in aspects of vagal activity, in experimental pain models; visceral pain (VP), neuropathy (CCI) and neurogenic infl ammation (NI).
Experimental procedures were performed on 130 Wistar rats in Pathophysiology Department of the Collegium Medicum, Jagiellonian University in Kraków. All the procedures were conducted with the approval of the I Local Ethical Committee for the Animal Studies in Kraków.
In this study we analyzed modifi cation of vagus nerve activity (electrical
activity) and c-Fos expression in vagal nodose ganglion (NG) after PTX administration. To estimate the intensity of pain symptoms manifestations we provided the behavioral tests adequate to the pain model: abdominal constriction test (WT) in VP, von Frey test to assess mechanical allodynia and Hargreaves test to assess thermal hyperalgesia in CCI and NI.
Visceral Pain (VP). There was used accepted animal model of the viscero-somatic pain [28, 29]. By injecting i.p. 0,5% acetic acid (10 ml/kg) there was induced chemical peritonitis.
Neuropathic pain (CCI). In the experiment there was used Bennet and
Xie model of chronic constriction injury (CCI) [30]. The behavioral tests were performed before the surgical procedure (day 0) and then on 14th day after the procedure. In our experiments the control group was the one, in which there was conducted the surgical procedure but without injuring the nerve itself (sham), additionally the healthy leg (contralateral, contra) — in every subgroup served as a control. These was evaluated and compared to the group with “pure” neuropathic pain (CCI) as the behavioral response of the injured leg (ipsilateral — ipsi).
Neurogenic Infl ammation (NI). Approved Gilchrist’s model was used for
induction of NI. Mechanical and thermal stimuli were administered 3 mm proximally to capsaicin injection place (highest pain response zone) [31].
Behavioral response in acute visceral pain — Writhing Test (WT). Intraperitoneal administration of acetic acid causes peritonitis and behavioral manifestation of nociception. The intensity of nociception was measured by counting of the rat’s abdominal and hind limb muscles contractions (WT) in a period of 30 minutes after acetic acid administration.
Von Frey’s test — test of mechanical allodynia. The von Frey’s test is used
to evaluation of the behavioral response to mechanical nociceptive stimuli [32, 33]. The experimental procedures were conducted by using automatic von Frey’s fi lament (Dynamic Plantar Aesthesiometer — Ugo Basile, Italy).
Hargreaves’s Test — test of thermal hyperalgesia. The Hargreaves’ test is
used to evaluation of the sensitivity to the thermal nociceptive stimuli [34]. To the tests there was used Plantar Test apparatus (Ugo Basile, Italy).
E l e c t r i c a l a c t i v i t y o f t h e l e f t v a g u s n e r v e
Under general anesthesia the left vagus nerve was exposed on the neck and recorded because it contains more afferent fi bers [35]. Furthermore in the trials conducted in the Pathophysiology Department there weren’t observed any differences between the left and right vagus nerve activity [36]. The nerve trunk was dissected transversely. On the distal part cuff electrode was placed. The signal was amplifi ed and converted by using the standard set (PowerLab/8SP ADInstruments Australia and AC/DC Differential Amplifi er A-M Systems USA). The records were archived and then worked up by using packet of software Chart 5 Pro ver. 5.4.2 extended with Spike Histogram v.1.2 (ADInstruments, Australia), compatible with Windows XP system. All the results were given as the mean ± standard deviation and statistically worked out. For the signifi cant accepted p values were p < 0,05.
c - F o s e x p r e s s i o n i n n o d o s e g a n g l i o n ( N G )
After Vetbutal overdosing inferior vagal ganglions (NG, nodose ganglions) were bilaterally prepared and sampled to histochemical evaluations. The prepara-tions were fi xed and then frozen. Frozen preparation was sliced by using cryo-stat and subsequently submitted to the immunohistochemical staining. The specimens were incubated with rabbit c-Fos antibody (K-25; Sc-253) (Santa Cruz Biotechnology). After 3 times repeated PBS washing the specimens were incubated with second degree biotinylated goat anti-rabbit antibody (Jack-son ImmunoResearch, West Grave, PA). Afterwards the specimens were in-cubated with streptavidin — Cy3 complex (Jackson ImmunoResearch, West Grave, PA). Then the specimens were embedded in fl uorescent specimen’s me-dium (DAKOCytomation, Denmark). Negative controls were conducted without the fi rst antibody. The specimens were analyzed under the Zeiss Axsioscop fl uorescent microscope.
P T X p r e - e m p t i v e a n a l g e s y
To the rats in the proper groups there was pre — emptively administered i.p. PTX in a dosage 10 mg/kg b.m. In the VP and NI groups PTX was given 15 minutes before conducting the behavioral tests. In the CCI group PTXwas administered every 12 hours during the experimental duration time — fi rst dosage the animals received 6 hours prior to the surgery. The aim was to limit the infl ammatory response in the evaluated groups.
The calculations were performed with using STATISTICA 8.0 for Windows
packet (StatSoft Inc., Tulusa, Oklahoma, USA). The results were given in the tabular and graphic form with using Microsoft Offi ce Excel 2007 software. Conformity of the quantitative variables distribution with the normal distribution was evaluated with using Shapiro–Wilk test. The results obtained from the analysis were given as the arithmetical mean ± standard deviation. As the statistically signifi cant results there were accepted those with the difference on the level p < 0,05. Graphs were made as deviation from control group (“pure” pain model).
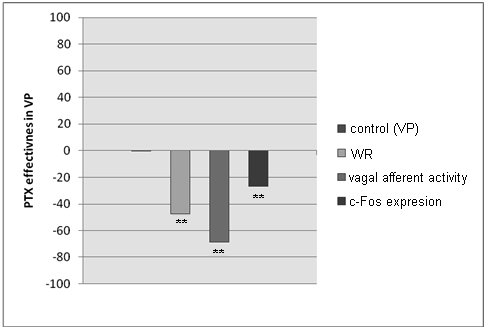
V i s c e r a l p a i n (VP). In model of VP we monitored the direct participation of the vagus nerve in mechanisms of visceral nociception which dominated over classic spinal pain pathways. PTX administration before experiment in visceral nociception decreased the pain response. Intra-peritoneal acetic acid administration increased both the vagal afferent discharges frequency and c-Fos expression in NG whereas pre-PTX administration suppressed this effect (Fig. 1, Tab. 1).
Fig. 1. Pain response (WR) and vagal activity after PTX administration in visceral pain.
Results are shown as deviation from VP group (0%) (** — p < 0.01)
Pain response (WR) and vagal activity after PTX administration in visceral pain
WR — number of writhes/30 min, Vagal Afferent Activity — frequency of impulses (Hz),
c-Fos Expression — number of c-Fos positive cells/slide of NG
N e u r o p a t h i c p a i n (CCI). The vagus nerve plays an important role in sensitization trough the development of chronic pain (CCI). In model of chronic pain (CCI) the vagus nerve is participating indirectly in both of peripheral and central sensitisation. The pain threshold signifi cantly increased after PTX. It allows to assume, that on the development of mechanical allodynia and of thermal hyperalgesia contribution of the vagus nerve is essential. Vagal afferents activity and c-fos expression in NG neurons in the CCI group increased. In the PTX group the vagus nerve activity and the c-Fos expression were lower in comparison to CCI (Fig. 2, Tab. 2), it demonstrate central modifi cation of vagal components.
Fig. 2. Behavioral Tests and vagal activity after PTX administration in CCI model.
Results are shown as deviation from “pure” CCI group (0) (* — < 0.05, ** — p < 0.01)
Behavioral Tests and vagal activity after PTX administration in CCI model
Von Frey’s Test — fi lament pressure (g), Hargreave’s Test — time of paw withdrawal (s),
Vagal Afferent Activity — frequency of impulses (Hz), c-Fos Expression — number of c-Fos
N e u r o g e n i c i n f l a m m a t i o n (NI). We observed attenuation of vagal
afferent activity in the left vagus nerve and absence of c-Fos expression in NG in the NI group. PTX didn’t infl uence the activity of the vagus nerve (Fig. 3, Tab. 3). These results support hypothesis of blockade of the central vagal centers in order to facilitate the nociceptive information transmitted by spinal pathways.
Fig. 3. Behavioral Tests and vagal activity after PTX administration in NI model.
Results are shown as deviation from “pure” NI group (0) (* — p < 0.05)
Behavioral Tests and vagal activity after PTX administration in NI model
Von Frey’s Test — fi lament pressure (g), Hargreave’s Test — time of paw withdrawal (s),
Vagal Afferent Activity — frequency of impulses (Hz), c-Fos Expression — number of c-Fos
PTX has strong antyinflamatory effects, decreases TNF-alpha and other proinfl ammatory cytokines (Il-1, Il-6, Il-8) production [37, 38]. In VP and CCI models we observed signifi cant increase in the pain threshold after blocking proinfl ammatory cytokines (PTX) whereas in NI there was no such effect [39]. Development of the local infl ammatory process activates the cells of the immune system, which release numerous infl ammatory mediators, such as cytokines, leukotrienes, prostaglandins, proteases, kinins, histamine. These mediators activate the sensory neurons or/and change their sensitivity [40–43]. The infl ammatory process gives the way to hyperalgesia that is documented by the animal experimental studies [44, 45]. In our studies we observed the increased electrical activity of the left vagus nerve in VP and CCI, on the contrary to NI model. Vagal afferent fi bers, due to the expression of the proinfl ammatory cytokines receptors, have a special place in the communication between immune system and CNS [46, 47]. Most of the authors describes the vagus nerve as responsible mostly for leading the physiological stimuli [48], however there are several data confi rming its role in pathophysiology of the nociceptive processes [4, 49, 50]. Pain stimulation after tissue damage and infl ammation modifi es nervous system response in important processes of peripheral and central sensitization. Vagus nerve plays a crucial role in the development of the autonomic anti–infl ammatory refl exes [51, 52]. This confi rms the Watkins et al. data, stating that post–infl ammatory hyperalgesia depends on the vagal afferent pathways integrity [53]. PTX was more effective in reduce of vagal afferents discharge in VP than in CCI model and it wasn’t effective in NI model. We also observed that PTX administration signifi cantly reduced c-Fos expression in nodose ganglon in VP, and even stronger in CCI model. In CCI model c-Fos expression in Dorsal Root Ganglia (DRG), spinal cord [54, 55], and in NTS [56, 57] increased. Complete dissection of the sciatic nerve also increased c-Fos expression in spinal cord [58] that with reference to our studies may prove the central sensitization, which may be mediated via vagal pathway. However
in antigen-induced arthritis (AIA) the c-Fos expression decreased in spinal cord [59]. According to Zhao et al. pain response to mechanical stimuli and to increased temperature in neuropathy doesn’t depend on the sympathetic nervous system [60]. There exists no marked evidence concerning the vagal participation in the acute peripheral pain models. Data obtained in our studies show that vagal afferent endings plays the crucial role in the chemical peritonitis model (VP). We observed increased vagal afferents activity and c-Fos expression in NG. Our fi nding is consistent with the others studies, reporting increase of gastric vagal afferents discharge and c-Fos expression in NTS induced by the different types of noxious stimuli [61, 62]. Holzer et al. show that exposure of the rat gastric mucosa to noxious stimuli is signaled to the brainstem, but not spinal cord. This observation is complemented by the fi nding that the visceromotor response to gastric acid challenge is suppressed by vagotomy, but not splanchnectomy [61, 63]. In other studies after intraperitoneal injection of IL-1β and LPS increased c-Fos expression in NTS and NG also confi rms our results [64, 65]. The involvement of the central modulation of spinal antinociceptive systems with the vagus nerve was observed in the rectum stretching model [66]. The participation of the vagal component in the chronic pain doesn’t limit itself only to the central nociception modulation. In the CCI model we observed complex, coming probably from the existence of “neuronal plasticity”, vagal participation in the modulation, both peripheral and central sensitization [5, 6]. These observations suggest that the vagus nerve may infl uence nociceptive information on many levels of its transmission, being in a close correlation with the immune and endocrine system especially at CNS level. In NI model we observed also the slight infl uence of the pre-emptively administered PTX on thermal hyperalgesia that could be a result of the acute character of the study and a lack of proper time for complete development of central sensitization [67]. Similar conclusions are coming from Obara et al. studies, where there was observed in CCI bigger infl uence of the infl ammatory component on the pain threshold increase than in NI [68]. This may confi rm the crucial role of the classic, nociceptive pathway in this model. Our results establish the participation of infl ammatory and vagal component in nociception what can constitute the base for using this tracks in both acute and chronic pain complexes treatment. Anti-infl ammatory substances (like PTX), by decreasing spinal and vagal activity, may modify peripheral and central sensitization. The vagus nerve is directly involved in the visceral nociceptive mechanisms, showing the predominance over the classical, spinal nociceptive pathways. Its activity depends on the infl ammatory component — so, it shows also antinociceptive action, dependent on the efferent pathways of the infl ammatory refl ex. In the neuropathic pain model vagus nerve is indirectly involved, playing its role in the peripheral and central sensitization mechanisms. The vagus nerve may modulate the higher CNS centers that are taking part in the nociception (classical
antinociceptive systems, endocrine system (HPA axis)). There is observed the central modulation of the vagal system through the spinal component that is playing the crucial role in the peripheral nociception. Pre-emptive blockage of the proinfl ammatory cytokines diminishes the development of the neuropathic signs. Suppression of pro-infl ammatory mediators synthesis in NI attenuates pain response not signifi cantly what can result from the used type of acute pain model and insuffi cient time needed for the full sensitization development.
In summary, our study demonstrates that preemptive inhibition of pro-
infl ammatory cytokine synthesis by treatment with PTX is useful in antago-nizing hyperalgesia in infl ammatory pain. Pentoxifylline reduces central and peri pheral sensitization processes depend on the vagal component in both acute and chronic pain models but in a different manner and mechanisms. Our results establish the participation of infl ammatory and vagal component in nociception. The modulation of the vagal system offers the new possibilities of the pain treatment in patients resistant to the classical analgesic therapy. 1. Bohotin C., Scholsem M., Bohotin V., Franzen R., Schoenen J.: Vagus nerve stimulation
attenuates heat- and formalin-induced pain in rats. Neurosci Lett 2003, 351(2): 79–82. —2.Bor- c kardt J.J., Kozel F.A., Anderson B., Walker A., George M.S.: Vagus nerve stimulation affects pain perception in depressed adults. Pain Res Manag. 2005; 10(1): 9–14. —3.Sedan O., Sprecher E., Yarnitsky D.: Vagal stomach afferents inhibit somatic pain perception. Pain. 2005, 113(3): 354–359. —4.Ciesielczyk K., Nowak Ł., Żurowski D., Thor P.J.: Vagal modulation of nociception in expe- rimental model of visceral pain. Folia Med Cracov. 2008, 49(3–4): 79–89. —5.Gebhart G.F.: Pathobiology of visceral pain: molecular mechanisms and therapeutic implications IV. Visceral afferent contributions to the pathobiology of visceral pain. Am J Physiol Gastrointest Liver Physiol. 2000, 278(6): G834–838. —6.Al-Chaer E.D., Traub R.J.: Biological basis of visceral pain: recent developments. Pain. 2002, 96(3): 221–225. —7.Randich A., Gebhart G.F.: Vagal afferent modu- lation of nociception. Brain Res Brain Res Rev. 1992, 17(2): 77–99. —8. Janig W., Khasar S.G., Levine J.D., Miao F.J.: The role of vagal visceral afferents in the control of nociception. Prog Brain Res. 2000, 122: 273–287. —9.Miao F.J., Janig W., Levine J.D.: Vagal branches involved in inhibition of bradykinin-induced synovial plasma extravasation by intrathecal nicotine and noxious stimulation in the rat. J Physiol. 1997, 498 (Pt 2): 473–481. —10.Miao F.J., Janig W., Dallman M.F., Benowitz N.L., Heller P.H., Basbaum A.I., et al.: Role of vagal afferents and spinal pathways modulating inhibition of bradykinin-induced plasma extravasation by intrathecal ni- cotine. J Neurophysiol. 1994, 72(3): 1199–1207. 11. Khasar S.G., Miao J.P., Janig W., Levine J.D.: Modulation of bradykinin-induced me-
chanical hyperalgesia in the rat by activity in abdominal vagal afferents. Eur J Neurosci. 1998,
10(2): 435–444. —12.Tien D., Ohara P.T., Larson A.A., Jasmin L.: Vagal afferents are necessary for the establishment but not the maintenance of kainic acid-induced hyperalgesia in mice. Pain. 2003, 102(1–2): 39–49. —13.Khasar S.G., Isenberg W.M., Miao F.J., Gear R.W., Green P.G., Le- vine J.D.: Gender and gonadal hormone effects on vagal modulation of tonic nociception. J Pain. 2001, 2(2): 91–100. —14.Maixner W., Dubner R., Kenshalo D.R., Jr., Bushnell M.C., Oliveras J.L.: Responses of monkey medullary dorsal horn neurons during the detection of noxious heat sti- muli. J Neurophysiol. 1989, 62(2): 437–449. —15.Maixner W., Bossut D.F., Whitsel E.A.: Eva- luation of vagal afferent modulation of the digastric refl ex in cats. Brain Res. 1991, 560(1–2): 55–62. —16.Randich A., Aicher S.A.: Medullary substrates mediating antinociception produced by electrical stimulation of the vagus. Brain Res. 1988, 445(1): 68–76. —17.Ren K., Randich A., Gebhart G.F.: Spinal serotonergic and kappa opioid receptors mediate facilitation of the tail fl ick refl ex produced by vagal afferent stimulation. Pain. 1991, 45(3): 321–329. —18.Ren K., Zhuo M., Randich A., Gebhart G.F.: Vagal afferent stimulation-produced effects on nociception in capsaicin- treated rats. J Neurophysiol. 1993, 69(5): 1530–1540. —19.Accetto B.: Benefi cial hemorheologic therapy of chronic peripheral arterial disorders with pentoxifylline: results of double-blind study versus vasodilator-nylidrin. Am Heart J. 1982, 103(5): 864–869. —20.Strieter R.M., Remick D.G., Ward P.A., Spengler R.N., Lynch J.P., 3rd, Larrick J., et al.: Cellular and molecular regulation of tumor necrosis factor-alpha production by pentoxifylline. Biochem Biophys Res Commun. 1988, 155(3): 1230–1236. 21. Zabel P., Schade F.U., Schlaak M.: Inhibition of endogenous TNF formation by pentoxifylline.
Immunobiology. 1993, 187(3–5): 447–463. —22.Dorazil-Dudzik M., Mika J., Schafer M.K., Li Y., Obara I., Wordliczek J., et al.: The effects of local pentoxifylline and propentofylline treatment on formalin-induced pain and tumor necrosis factor-alpha messenger RNA levels in the infl amed tissue of the rat paw. Anesth Analg. 2004, 98(6): 1566–1573, table of contents. —23.Lu C.H., Chao P.C., Borel C.O., Yang C.P., Yeh C.C., Wong C.S., et al.: Preincisional intravenous pentoxifyl- line attenuating perioperative cytokine response, reducing morphine consumption, and improving recovery of bowel function in patients undergoing colorectal cancer surgery. Anesth Analg. 2004, 99(5): 1465–1471; table of contents. —24.Ji Q., Zhang L., Jia H., Xu J.: Pentoxifylline inhibits endotoxin-induced NF-kappa B activation and associated production of proinfl ammatory cytokines. Ann Clin Lab Sci. 2004, 34(4): 427–436. —25.Vale M.L., Benevides V.M., Sachs D., Brito G.A., da Rocha F.A., Poole S., et al.: Antihyperalgesic effect of pentoxifylline on experimental infl ammatory pain. Br J Pharmacol. 2004, 143(7): 833–844. —26.Liu J., Feng X., Yu M., Xie W., Zhao X., Li W., et al.: Pentoxifylline attenuates the development of hyperalgesia in a rat model of neuropathic pain. Neurosci Lett 2007, 412(3): 268–272. —27.Nowak L., Żurowski D., Garlicki J., Thor P.J.: [Role of pendoxifi line (PTX) in different and cute and chronic models of pain in rats]. Folia Med Cracov. 2008, 49(3–4): 103–109. —28.Nakamura H., Shimizu M.: Site of analgesic action of a non-steroidal, anti-infl ammatory drug, tolmetin sodium, in rats. Br J Pharmacol. 1981, 73(3): 779–785. —29.Wang H.C., Chai S.C., Wu Y.S., Wang C.C.: Does the medial thalamus play a role in the negative affective component of visceral pain in rats? Neurosci Lett. 2007, 420(1): 80–84. —30.Bennett G.J., Xie Y.K.: A peripheral mononeuropathy in rat that produces disorders of pain sensation like those seen in man. Pain. 1988, 33(1): 87–107. 31. Gilchrist H.D., Allard B.L., Simone D.A.: Enhanced withdrawal responses to heat and me-
chanical stimuli following intraplantar injection of capsaicin in rats. Pain. 1996, 67(1): 179–188. —32.Villetti G., Bergamaschi M., Bassani F., Bolzoni P.T., Maiorino M., Pietra C., et al.: Antinoci- ceptive activity of the N-methyl-D-aspartate receptor antagonist N-(2-Indanyl)-glycinamide hydro- chloride (CHF3381) in experimental models of infl ammatory and neuropathic pain. J Pharmacol Exp Ther. 2003, 306(2): 804–814. —33.Tal M., Bennett G.J.: Extra-territorial pain in rats with a peripheral mononeuropathy: mechano-hyperalgesia and mechano-allodynia in the territory of an uninjured nerve. Pain. 1994, 57(3): 375–382. —34. Hargreaves K., Dubner R., Brown F., Flores C., Joris J.: A new and sensitive method for measuring thermal nociception in cutaneous
hyperalgesia. Pain. 1988, 32(1): 77–88. —35.Berthoud H.R., Patterson L.M., Neumann F., Neu- huber W.L.: Distribution and structure of vagal afferent intraganglionic laminar endings (IGLEs) in the rat gastrointestinal tract. Anat Embryol (Berl). 1997, 195(2): 183–191. —36.Krolczyk G., Żurowski D., Sobocki J., Laskiewicz J., Thor P.J.: Encoding meal in integrated vagal afferent dis- charge. J Physiol Pharmacol. 2004, 55(1 Pt 1): 99–106. —37.El-Ghoneimi A., Cursio R., Schmid- -Alliana A., Tovey M., Lasfar A., Michiels J.F., et al.: Pentoxifylline inhibits liver expression of tumor necrosis factor alpha mRNA following normothermic ischemia-reperfusion. HPB (Oxford) 2007, 9(2): 112–119. —38.Selim K., Huseyin C., Ibrahim K.H., Hasan B.U., Kazim U., Huseyin K.: Effect of pentoxifylline on tumor necrosis factor-alpha and interleukin-6 levels in neonatal sepsis. Med J Malaysia. 2004, 59(3): 391–394. —39.Saade N.E., Jabbur S.J.: Nociceptive behavior in animal models for peripheral neuropathy: spinal and supraspinal mechanisms. Prog Neurobiol. 2008, 86(1): 22–47. —40.Bueno L., Fioramonti J.: Visceral perception: infl ammatory and non- -infl ammatory mediators. Gut. 2002, 51 Suppl 1: i19–23. 41. Huang J., Zhang N.C., Zhou J., Yang Z.H.: [Effects of stimulation of intact vagus nerve
on systemic infl ammatory response in rats]. Zhonghua Shao Shang Za Zhi. 2008, 24(2): 99–101. —42.Rogers R.C., Van Meter M.J., Hermann G.E.: Tumor necrosis factor potentiates central vagal afferent signaling by modulating ryanodine channels. J Neurosci. 2006, 26(49): 12642–12646. —43.Sengupta J.N., Saha J.K., Goyal R.K.: Differential sensitivity to bradykinin of esophageal distension-sensitive mechanoreceptors in vagal and sympathetic afferents of the opossum. J Neu- rophysiol. 1992, 68(4): 1053–1067. —44.Lamb K., Kang Y.M., Gebhart G.F., Bielefeldt K.: Nerve growth factor and gastric hyperalgesia in the rat. Neurogastroenterol Motil. 2003, 15(4): 355–361. —45.Ozaki N., Bielefeldt K., Sengupta J.N., Gebhart G.F.: Models of gastric hyperalgesia in the rat. Am J Physiol Gastrointest Liver Physiol. 2002, 283(3): G666–676. —46.Danzer M., Jocic M., Samberger C., Painsipp E., Bock E., Pabst M.A., et al.: Stomach-brain communication by vagal afferents in response to luminal acid backdiffusion, gastrin, and gastric acid secretion. Am J Phy- siol Gastrointest Liver Physiol. 2004, 286(3): G403–411. —47.Lamb K., Kang Y.M., Gebhart G.F., Bielefeldt K.: Gastric infl ammation triggers hypersensitivity to acid in awake rats. Gastroentero- logy. 2003, 125(5): 1410–1418. —48.Grundy D.: Neuroanatomy of visceral nociception: vagal and splanchnic afferent. Gut. 2002, 51 Suppl 1: i2–5. —49.Weissman-Fogel I., Dashkovsky A., Rogowski Z., Yarnitsky D.: Vagal damage enhances polyneuropathy pain: additive effect of two algogenic mechanisms. Pain. 2008, 138(1): 153–162. —50.Żurowski D., Nowak L., Thor P.J.: [The role of vagal afferents in visceral hyperalgesia]. Folia Med Cracov. 2005, 46(3–4): 97–105. 51. Pavlov V.A., Tracey K.J.: Controlling infl ammation: the cholinergic anti-infl ammatory
pathway. Biochem Soc Trans. 2006, 34(Pt 6): 1037–1040. —52.Tracey K.J.: The infl ammatory refl ex. Nature. 2002, 420(6917): 853–859. —53.Watkins L.R., Wiertelak E.P., Goehler L.E., Mooney- -Hei berger K., Martinez J., Furness L., et al.: Neurocircuitry of illness-induced hyperalgesia. Brain Res. 1994, 639(2): 283–299. —54.Morgado C., Tavares I.: C-fos expression at the spinal dorsal horn of streptozotocin-induced diabetic rats. Diabetes Metab Res Rev. 2007, 23(8): 644–652. —55.Walczak J.S., Pichette V., Leblond F., Desbiens K., Beaulieu P.: Characterization of chronic constriction of the saphenous nerve, a model of neuropathic pain in mice showing rapid molecu- lar and electrophysiological changes. J Neurosci Res. 2006, 83(7): 1310–1322. —56.Gamboa- -Esteves F.O., McWilliam P.N., Batten T.F.: Substance P (NK1) and somatostatin (sst2A) receptor immunoreactivity in NTS-projecting rat dorsal horn neurones activated by nociceptive afferent in- put. J Chem Neuroanat. 2004, 27(4): 251–266. —57.Haxhiu M.A., Strohl K.P., Cherniack N.S.: The N-methyl-D-aspartate receptor pathway is involved in hypoxia-induced c-Fos protein expression in the rat nucleus of the solitary tract. J Auton Nerv Syst. 1995, 55(1–2): 65–68. —58.Chi S.I., Levine J.D., Basbaum A.I.: Peripheral and central contributions to the persistent expression of spinal cord fos-like immunoreactivity produced by sciatic nerve transection in the rat. Brain Res. 1993, 617(2): 225–237. —59.Abbadie C., Besson J.M., Calvino B.: c-Fos expression in the spinal cord and pain-related symptoms induced by chronic arthritis in the rat are prevented by pre-
treatment with Freund adjuvant. J Neurosci. 1994, 14(10): 5865–5871. —60.Zhao C., Chen L., Tao Y.X., Tall J.M., Borzan J., Ringkamp M., et al.: Lumbar sympathectomy attenuates cold allo- dynia but not mechanical allodynia and hyperalgesia in rats with spared nerve injury. J Pain. 2007, 8(12): 931–937. 61. Holzer P.: Afferent signalling of gastric acid challenge. J Physiol Pharmacol. 2003, 54
Suppl 4: 43–53. —62.Kang Y.M., Bielefeldt K., Gebhart G.F.: Sensitization of mechanosens- itive gastric vagal afferent fi bers in the rat by thermal and chemical stimuli and gastric ulcers. J Neurophysiol. 2004, 91(5): 1981–1989. —63.Mertz H.: Review article: visceral hypersensitivity. Aliment Pharmacol Ther 2003, 17(5): 623–633. —64. Goehler L.E., Gaykema R.P., Hammack S.E., Maier S.F., Watkins L.R.: Interleukin-1 induces c-Fos immunoreactivity in primary afferent neu- rons of the vagus nerve. Brain Res. 1998, 804(2): 306–310. —65.Goehler L.E., Gaykema R.P., Hansen M.K., Anderson K., Maier S.F., Watkins L.R.: Vagal immune-to-brain communication: a vi- sceral chemosensory pathway. Auton Neurosci. 2000, 85(1–3): 49–59. —66.Gschossmann J.M., Mayer E.A., Miller J.C., Raybould H.E.: Subdiaphragmatic vagal afferent innervation in activation of an opioidergic antinociceptive system in response to colorectal distension in rats. Neuroga- stroenterol Motil. 2002, 14(4): 403–408. —67.Nowak Ł., Żurowski D., Garlicki J., Thor P.J.: Role of pentoxifi line in selected acute and chronic models of pain in rats. Folia Med Cracov. 2008, 49. —68.Obara I., Makuch W., Spetea M., Schutz J., Schmidhammer H., Przewlocki R., et al.: Local peripheral antinociceptive effects of 14-O-methyloxymorphone derivatives in infl ammatory and neuropathic pain in the rat. Eur J Pharmacol. 2007, 558(1–3): 60–67.
2 Department of Pain Treatment and Palliative Care
ul. Śniadeckich 10, 31-531 Kraków, Poland
3 Department of Pain Research and Treatment
ul. Śniadeckich 10, 31-531 Kraków, Poland
Corresponding author:
Estradiol and Exercise-Induced Creatine Kinase Activity JEP online Journal of Exercise Physiology online Official Journal of The American Society of Exercise Physiologists (ASEP) ISSN 1097-9751 Volume 4 Number 2 May 2001 EFFECTS OF CIRCULATING ESTRADIOL ON EXERCISE-INDUCED CREATINEKINASE ACTIVITYSTEPHEN M. ROTH1, RICHARD GAJDOSIK2 AND BRENT C. RUBY11 Human Performance Labora
C.CLIN Sud-Est – Septembre 2004 Conduite à tenir en cas d’une épidémie de gale en établissement de santé Définition : la gale est une ectoparasitose (parasite externe) à Sarcoptes scabiei variété hominis qui vit dans l’épiderme humain, à l’origine d’une dermatose très prurigineuse et contagieuse. I. Contexte épidémiologique Le sarcopte a un aspec
 The calculations were performed with using STATISTICA 8.0 for Windows
packet (StatSoft Inc., Tulusa, Oklahoma, USA). The results were given in the tabular and graphic form with using Microsoft Offi ce Excel 2007 software. Conformity of the quantitative variables distribution with the normal distribution was evaluated with using Shapiro–Wilk test. The results obtained from the analysis were given as the arithmetical mean ± standard deviation. As the statistically signifi cant results there were accepted those with the difference on the level p < 0,05. Graphs were made as deviation from control group (“pure” pain model).
V i s c e r a l p a i n (VP). In model of VP we monitored the direct participation of the vagus nerve in mechanisms of visceral nociception which dominated over classic spinal pain pathways. PTX administration before experiment in visceral nociception decreased the pain response. Intra-peritoneal acetic acid administration increased both the vagal afferent discharges frequency and c-Fos expression in NG whereas pre-PTX administration suppressed this effect (Fig. 1, Tab. 1).
Fig. 1. Pain response (WR) and vagal activity after PTX administration in visceral pain.
The calculations were performed with using STATISTICA 8.0 for Windows
packet (StatSoft Inc., Tulusa, Oklahoma, USA). The results were given in the tabular and graphic form with using Microsoft Offi ce Excel 2007 software. Conformity of the quantitative variables distribution with the normal distribution was evaluated with using Shapiro–Wilk test. The results obtained from the analysis were given as the arithmetical mean ± standard deviation. As the statistically signifi cant results there were accepted those with the difference on the level p < 0,05. Graphs were made as deviation from control group (“pure” pain model).
V i s c e r a l p a i n (VP). In model of VP we monitored the direct participation of the vagus nerve in mechanisms of visceral nociception which dominated over classic spinal pain pathways. PTX administration before experiment in visceral nociception decreased the pain response. Intra-peritoneal acetic acid administration increased both the vagal afferent discharges frequency and c-Fos expression in NG whereas pre-PTX administration suppressed this effect (Fig. 1, Tab. 1).
Fig. 1. Pain response (WR) and vagal activity after PTX administration in visceral pain.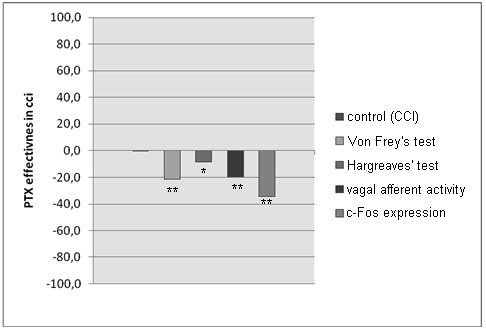 Pain response (WR) and vagal activity after PTX administration in visceral pain
WR — number of writhes/30 min, Vagal Afferent Activity — frequency of impulses (Hz),
c-Fos Expression — number of c-Fos positive cells/slide of NG
N e u r o p a t h i c p a i n (CCI). The vagus nerve plays an important role in sensitization trough the development of chronic pain (CCI). In model of chronic pain (CCI) the vagus nerve is participating indirectly in both of peripheral and central sensitisation. The pain threshold signifi cantly increased after PTX. It allows to assume, that on the development of mechanical allodynia and of thermal hyperalgesia contribution of the vagus nerve is essential. Vagal afferents activity and c-fos expression in NG neurons in the CCI group increased. In the PTX group the vagus nerve activity and the c-Fos expression were lower in comparison to CCI (Fig. 2, Tab. 2), it demonstrate central modifi cation of vagal components.
Fig. 2. Behavioral Tests and vagal activity after PTX administration in CCI model.
Pain response (WR) and vagal activity after PTX administration in visceral pain
WR — number of writhes/30 min, Vagal Afferent Activity — frequency of impulses (Hz),
c-Fos Expression — number of c-Fos positive cells/slide of NG
N e u r o p a t h i c p a i n (CCI). The vagus nerve plays an important role in sensitization trough the development of chronic pain (CCI). In model of chronic pain (CCI) the vagus nerve is participating indirectly in both of peripheral and central sensitisation. The pain threshold signifi cantly increased after PTX. It allows to assume, that on the development of mechanical allodynia and of thermal hyperalgesia contribution of the vagus nerve is essential. Vagal afferents activity and c-fos expression in NG neurons in the CCI group increased. In the PTX group the vagus nerve activity and the c-Fos expression were lower in comparison to CCI (Fig. 2, Tab. 2), it demonstrate central modifi cation of vagal components.
Fig. 2. Behavioral Tests and vagal activity after PTX administration in CCI model.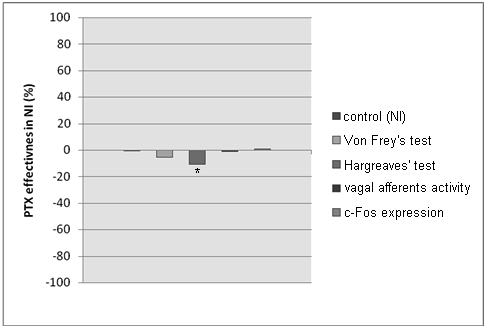 Behavioral Tests and vagal activity after PTX administration in CCI model
Von Frey’s Test — fi lament pressure (g), Hargreave’s Test — time of paw withdrawal (s),
Vagal Afferent Activity — frequency of impulses (Hz), c-Fos Expression — number of c-Fos
N e u r o g e n i c i n f l a m m a t i o n (NI). We observed attenuation of vagal
afferent activity in the left vagus nerve and absence of c-Fos expression in NG in the NI group. PTX didn’t infl uence the activity of the vagus nerve (Fig. 3, Tab. 3). These results support hypothesis of blockade of the central vagal centers in order to facilitate the nociceptive information transmitted by spinal pathways.
Fig. 3. Behavioral Tests and vagal activity after PTX administration in NI model.
Behavioral Tests and vagal activity after PTX administration in CCI model
Von Frey’s Test — fi lament pressure (g), Hargreave’s Test — time of paw withdrawal (s),
Vagal Afferent Activity — frequency of impulses (Hz), c-Fos Expression — number of c-Fos
N e u r o g e n i c i n f l a m m a t i o n (NI). We observed attenuation of vagal
afferent activity in the left vagus nerve and absence of c-Fos expression in NG in the NI group. PTX didn’t infl uence the activity of the vagus nerve (Fig. 3, Tab. 3). These results support hypothesis of blockade of the central vagal centers in order to facilitate the nociceptive information transmitted by spinal pathways.
Fig. 3. Behavioral Tests and vagal activity after PTX administration in NI model.