Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Untitled
Journal of Veterinary Emergency and Critical Care 17(1) 2007, pp 61–66
Comparison of the hypothalamic^pituitary^adrenalaxis in MDR1-1D and MDR1 wildtype dogs
Katrina L. Mealey, DVM, PhD, DACVIM, DACVCP, John M. Gay, DVM, PhD, DACVPM,Linda G. Martin, DVM, MS, DACVECC and Denise K. Waiting, LVT
Objective: To evaluate the hypothalamic–pituitary–adrenal (HPA) axis in MDR1-1D (dogs with the MDR1mutation associated with ivermectin sensitivity) and MDR1 wildtype dogs. Design: Prospective study. Setting: Institutional vivarium. Animals: Seven healthy Collie dogs. Measurements: MDR1 genotyping was used for allocation of dogs to 1 of 2 groups: dogs homozygous for thewildtype MDR1 allele (MDR1 wildtype) and those homozygous for the MDR1-1D mutation (MDR1 mutant). Blood samples were obtained for determination of cortisol and adrenocorticotropin hormone (ACTH)concentrations under basal conditions, before and after ACTH administration, and before and afterdexamethasone administration. Main results: Significant differences were identified between the MDR1 mutant and MDR1 wildtype groups. Basal plasma cortisol concentrations and cortisol concentrations after ACTH administration were significantlylower in MDR1 mutant dogs as compared with MDR1 wildtype dogs. Plasma ACTH concentrations afterdexamethasone administration were significantly lower in MDR1 mutant dogs as compared with MDR1wildtype dogs. Conclusions: Results suggest that P-glycoprotein (P-gp) plays a role in regulation of the HPA axis. Furthermore, it appears that the HPA axis in MDR1 mutant dogs that lack P-gp is suppressed compared withMDR1 wildtype dogs. This finding may explain some clinical observations in breeds known to harbor theMDR1 mutation including Collies, Shelties, Australian Shepherds, and others. There is a clinical impressionthat many of these dogs have worse outcomes in response to stress and, at times, respond poorly toappropriate therapy. HPA axis suppression, secondary to the MDR1 mutation, could result in a relativeadrenal insufficiency (RAI) state during times of stress or illness. Further studies are required to determine therelationship between the MDR1 genotype and RAI.
(J Vet Emerg Crit Care 2007; 17(1): 61–66) doi: 10.1111/j.1476-4431.2006.00196.x
Keywords: blood–brain barrier, collie, cortisol, P-glycoprotein, relative adrenal insufficiency
capillary lumen. P-gp appears to be highly homolo-gous, but not identical, among mammalian species.
P-glycoprotein (P-gp), the product of the MDR1 or
Drugs that are known substrates for canine P-gp in-
ABCB1 gene, is a crucial component of the blood–brain
clude ivermectin, loperamide, vincristine, vinblastine,
barrier, protecting the brain from many potentially toxic
and doxorubicin.2–5 Over 50 drugs have been shown to
xenobiotics.1 P-gp functions as an ATP-dependent drug
be substrates for human and murine P-gp6,7 and, be-
transporter that is expressed on the luminal membrane
cause of the high degree of homology of P-gp between
of brain capillary endothelial cells where it transports a
species, it is suspected that these same drugs are subst-
variety of substrates from the brain tissue back into the
rates for canine P-gp. Individuals that lack P-gp, asoccurs in herding breed dogs with the MDR1-1D
From the Department of Veterinary Clinical Sciences, College of VeterinaryMedicine, Washington State University, Pullman, WA.
mutation, are highly susceptible to neurotoxicosis afterroutine doses of P-gp substrate drugs such as ivermec-
Address correspondence and reprint requests to:Katrina L. Mealey, Department of Veterinary Clinical Sciences, College
tin and loperamide.8–11 This susceptibility underscores
of Veterinary Medicine, Washington State University, Pullman,
the importance of P-gp in limiting exposure of the brain
WA 99164-6610. E-mail: kmealey@vetmed.wsu.edu
& Veterinary Emergency and Critical Care Society 2006
Exogenous substances are not the only substrates for
P-gp. In rodents, endogenous hormones including cor-ticosterone are substrates for P-gp suggesting thatP-gp may have a role in regulating their plasma con-centrations.12 Results from recent studies support this
contention. In rodent studies, P-gp was shown to re-strict access of corticosterone and cortisol to the brain.12Furthermore, abcb1ab ( À / À ) double knockout micethat lack P-gp have a suppressed hypothalamic–pitu-itary–adrenal (HPA) axis compared with wildtype
mice.13 Collectively, these results suggest that P-gpnormally limits the concentration of cortisol and corti-
costerone at the hypothalamus and pituitary bluntingfeedback inhibition of the HPA axis. If this premise is
correct, then higher concentrations of cortisol and cor-ticosterone would be expected to reach the hypo-thalamus and pituitary in animals that lack P-gp re-sulting in greater feedback inhibition of the HPA axis
and low endogenous cortisol levels (Figure 1). Conse-
quently, dogs such as herding breed dogs with theMDR1-1D mutation would be expected to be predis-posed to relative adrenal insufficiency (RAI).
RAI is characterized by inadequate production of
cortisol in relation to an increased physiological de-
mand during periods of stress such as critical ill-
ness.14,15 Human patients with RAI have a reducedcapacity to cope with critical illness and these patients
Figure 1: Illustration of the hypothalamic–pituitary–adrenal
have been shown to have a poorer outcome than pa-
(HPA) axis in wildtype dogs (A) and MDR1 mutant dogs (B).
tients with a normal HPA axis.14 Interestingly, some
In wildtype dogs, P-glycoprotein (P-gp) is present at the blood–
veterinarians have described Collies as ‘wimpy’ or as
brain barrier and limits entry of cortisol into the brain further
‘not participating in their own recovery’ because some
limiting cortisol’s feedback inhibition of corticotrophin releasing
individuals of this breed have had poorer outcomes or
hormone (CRH) secretion and adrenocorticotropin hormone
have not responded as well as dogs of other breeds
(ACTH). In MDR1 mutant dogs, P-gp is not present at the
with similar illnesses.a These anecdotal observations
blood–brain barrier, therefore, greater concentrations of cortisol
are consistent with the hypothesis that dogs with the
are present within the brain enabling cortisol to exert feedback
MDR1-1D mutation have a blunted HPA axis compared
inhibition of CRH, ACTH, and, ultimately, cortisol secretion. Solid lines indicate stimulatory effects (with bolder lines indi-
with MDR1 wildtype dogs. Therefore, the purpose of
cating greater stimulation) while dashed lines indicate inhibi-
this study was to investigate HPA system regulation in
tory effects (with bolder lines indicating greater inhibition).
MDR1-1D and MDR1 wildtype dogs utilizing dexameth-asone suppression and adrenocorticotropin hormone
2 females) were included in the study. Genotyping
was performed at a commercial laboratory.b
For each dog, baseline ACTH values were measured on
All aspects of this study were approved by the Insti-
3 occasions and baseline cortisol values were measured
tutional Animal Care and Use Committee. Seven Collie
on 5 occasions. ACTH stimulation and dexamethasone
dogs were studied ranging in age from 1 to 8 years of
suppression studies were performed once. Studies were
age. Dogs were determined to be healthy on the basis of
performed such that collection of basal (prestimulation
results from physical examination, a complete blood
or presuppression) samples occurred at approximately
count, serum biochemistry panel, and urinalysis. Three
08:30 hours ( Æ 30 minutes). Blood was collected by
dogs that were homozygous for the MDR1 wildtype
jugular venipuncture. Blood was injected into EDTA
genotype (1 male, 2 females) and 4 dogs that were
tubes for cortisol determination. For ACTH determina-
homozygous for the MDR1-1D mutation (2 males,
tion, blood was injected into prechilled EDTA tubes
& Veterinary Emergency and Critical Care Society 2006, doi: 10.1111/j.1476-4431.2006.00196.x
with a protease inhibitor, 100 mL aprotinin, added. Plas-ma was harvested by centrifugation at 4 1C. Plasma wastransferred to prechilled cryovials and frozen at
À 80 1C until analyzed. Cortisol and ACTH concentra-
tions were determined by a commercial laboratoryc us-ing methodology validated for the dog as previouslydescribed.16 A washout period of at least 2 weeks wasallowed between each study.
ACTH stimulation testingACTH (1 mg/kg) was administered intravenously (IV)(cephalic vein). Blood was collected before (time 0) and1 hour after ACTH administration for determination ofplasma cortisol concentrations.
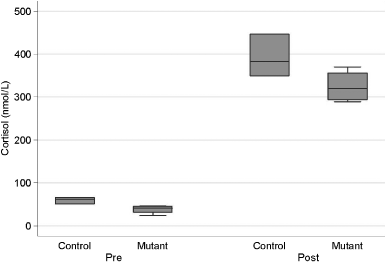
Figure 2: Box and whisker plot of basal plasma cortisol samples
in MDR1 mutant dogs (mutant; n 5 4 dogs, 3 samples per dog)
Three different doses of dexamethasone (0.05, 0.01, and
and MDR1 wildtype dogs (control; n 5 3 dogs, 3 samples per
0.001 mg/kg) were used. Dexamethasone was injected
dog). Basal plasma cortisol concentrations are significantly low-
IV (cephalic vein). Blood was collected before (time 0),
er in MDR1 mutant dogs (nP 5 0.044) as compared with MDR1
4, and 8 hours after dexamethasone injection for deter-
wildtype dogs. Box, interquartile range (IQR); line, median;
mination of cortisol and ACTH concentrations.
whiskers, 1.5 Â IQR; dots, data points greater than 1.5 Â IQR.
vs. 226 Æ 12.7 nmol/L SEM, F1, 5 5 7.02, P 5 0.045), but
Data were entered into a spreadsheetd and statistical
the degree of response (i.e., slope) was not different
analyses were performed using a commercial statistics
program.e Because of repeated sampling over fixed
For dexamethasone suppression testing, statistically
times after multiple levels of stimulation, repeated
significant differences in plasma cortisol concentrations
measures ANOVA models with nested subjects as ran-
between MDR1 mutant and wildtype dogs in response
dom factors were employed. Standardized residuals
to dexamethasone administration were not detected
were plotted to detect departures from normality.
at any dexamethasone dose (Figure 4). At the 4- and
Greenhouse–Geisser epsilon values were calculated
8-hour time points after 0.05 mg/kg of dexamethasone
for the repeated measures to determine if F-test de-grees of freedom required reduction for deviations fromcompound symmetry and were adjusted accordingly. Acritical value of 0.05 was used as the threshold of sta-tistical significance and results are presented withstandard error of the mean (SEM) as an indication ofdata variability.
For each dog, baseline ACTH values were measured on3 occasions and baseline cortisol values on 5 occasions. A statistically significant association was observed be-tween
P 5 0.044), but not for ACTH (P 5 0.24). Mean basalcortisol concentrations in MDR1 mutant dogs were
Figure 3: Box and whisker plot of plasma cortisol concentra-
39.3 Æ 6.18 versus 61.2 Æ 5.35 nmol/L SEM for MDR1
tions in MDR1 wildtype dogs (control; n 5 3 dogs) and MDR1
wildtype dogs. Figure 2 shows a box and whisker plot
mutant dogs (mutant; n 5 3 dogs) immediately before (pre) and
of basal cortisol concentrations in MDR1 mutant and
1 hour after (post)-intravenous administration of 1 mg/kg syn-
MDR1 wildtype dogs. For ACTH stimulation testing,
thetic adrenocorticotropin hormone (ACTH). The mean plasma
MDR1 mutant dogs displayed significantly lower over-
cortisol concentration after ACTH administration was signifi-
all mean plasma cortisol concentrations after ACTH
cantly lower in MDR1 mutant dogs (n) than in MDR1 wildtype
administration than MDR1 wildtype dogs (181.5 Æ 11.0
& Veterinary Emergency and Critical Care Society 2006, doi: 10.1111/j.1476-4431.2006.00196.x
Cortisol Concentration (nmol 10 ACTH Concentration (pg Time (hours) Time (hours)
Figure 4: Mean (SEM) plasma cortisol concentrations in MDR1wildtype dogs (wild; n 5 3) and MDR1 mutant dogs (mutant;
Figure 5: Mean (SEM) plasma adrenocorticotropin hormone
n 5 4) before (time 0), 4, and 8 hours after dexamethasone ad-
(ACTH) concentrations in MDR1 wildtype dogs (wild; n 5 3)
ministration. Dexamethasone was administered at 3 different
and MDR1 mutant dogs (mutant; n 5 4) before (time 0), 4, and 8
doses with at least a 2-week washout period between dosing.
hours after dexamethasone administration. A significant inter-
Statistically significant differences in plasma cortisol concentra-
action (P 5 0.037) was detected between genotype (MDR1 sta-
tions between MDR1 mutant and wildtype dogs in response to
tus) and ACTH plasma concentrations. Dexamethasone was
dexamethasone administration were not detected.
administered at 2 different doses with at least a 2-week washoutperiod between dosing.
was administered, plasma ACTH concentrations fellbelow the limit of quantitation and so statistical com-parisons were not performed. However, a significant
tion by allowing higher brain concentrations of endog-
between genotype (MDR1 status) and ACTH plasma
releasing hormone (CRH) secretion. Studies were per-
concentrations. Means for the MDR1 mutant dogs
formed in the abcb1ab double knockout mice to test this
receiving 0.01 mg/kg dexamethasone dose were signif-
hypothesis. Compared with wildtype mice, abcb1ab
icantly lower (12.67 Æ 1.47 pg/mL SEM) than MDR1
( À / À ) double knockout mice have lower plasma cor-
wildtype dogs (22.89 Æ 1.70 pg/mL SEM) receiving that
tisol concentrations under basal conditions and under
dose of dexamethasone (Figure 5). At the lowest dexa-
conditions of stress.13 The knockout mice also have
methasone dose (0.001 mg/kg), a statistically significant
downregulated CRH mRNA expression in the hypo-
difference in plasma ACTH concentrations was not
thalamic paraventricular nucleus compared with wild-
type mice suggesting a sustained suppression of theHPA system.13 Because MDR1 mutant dogs are pheno-typically similar to abcb1ab ( À / À ) double knockout
mice with respect to brain penetration of other P-gp
P-gp functions as an ATP-dependent drug efflux pump
substrates (e.g., ivermectin, loperamide), it was sus-
that is capable of transporting substrate drugs against a
pected that MDR1 mutant dogs might show evidence of
concentration gradient from the intracellular to the ex-
tracellular space.17 It is expressed on the luminal aspect
The present data suggest that lack of P-gp function at
of brain capillary endothelial cells where it functions to
the blood–brain barrier in MDR1 mutant dogs results in
limit brain penetration of substrate xenobiotics includ-
altered activity and regulation of the HPA axis. Similar
ing the endogenous hormones, corticosterone, and co-
to results from rodent studies, mean basal cortisol con-
rtisol.1,17,18 Studies in rodents have shown that P-gp
centrations in MDR1 mutant dogs (39.3 Æ 6.18 nmol/L
limits access of corticosterone to the brain.12 Further
SEM) were significantly lower than in MDR1 wildtype
studies were performed in genetically engineered
dogs (61.2 Æ 5.35 nmol/L). It is presumed that this ap-
abcb1ab double knockout mice that are phenotypically
parent suppression of systemic cortisol concentrations
similar to MDR1 mutant dogs since both lack P-gp. Be-
in MDR1 mutant dogs results from greater cortisol
cause P-gp normally limits access of endogenous cor-
feedback on hypothalamic paraventricular neurons in
ticosteroids to the brain, it was hypothesized that lack
these dogs as compared with wildtype dogs that have
of P-gp might profoundly influence HPA axis regula-
normal P-gp function. The fact that MDR1 mutant dogs
& Veterinary Emergency and Critical Care Society 2006, doi: 10.1111/j.1476-4431.2006.00196.x
had lower plasma cortisol concentrations, even after
as acute illness, resulting in RAI or an RAI-like
ACTH stimulation, suggests that there is chronic sup-
pression of cortisol production. This observation raises
RAI is a syndrome characterized by insufficient pro-
questions as to the ability of MDR1 mutant dogs to
duction of cortisol in relation to an increased demand
during periods of severe stress, particularly in critical
Results of dexamethasone suppression tests from the
illnesses such as sepsis or septic shock.14,19 The syn-
present study differed somewhat from results reported
drome is presumed to be associated with altered func-
for abcb1ab ( À / À ) double knockout mice. In both
tion of the HPA axis. Insufficient stress hormone
MDR1 mutant dogs and abcb1ab ( À / À ) double knock-
synthesis appears to be a transient phenomenon in hu-
out mice, plasma concentrations of ACTH were signif-
man RAI patients since life-long replacement of cor-
icantly lower than in their wildtype counterparts.
ticosteroids (as would be essential in patients with true
However, while plasma concentrations of cortisol were
hypoadrenocorticism) is not necessary.19 From a clinical
suppressed at lower dexamethasone doses in abcb1ab
perspective, it is extremely important to recognize sep-
( À / À ) double knockout mice as compared with wild-
tic patients with RAI because these patients tend to
type mice, there was not a statistically significant dif-
carry a worse outcome if untreated. Treatment, which
consists of low (‘physiologic’) doses of corticosteroids,
dexamethasone administration in MDR1 mutant versus
appears to reduce morbidity and mortality rates, par-
ticularly in septic patients.20 Although RAI is well doc-
It is not clear why ACTH concentrations following
umented in critically ill human patients,20–22 relatively
dexamethasone suppression differ in MDR1 mutant
less is known about adrenal dysfunction in critically ill
dogs as compared with wildtype dogs while cortisol
veterinary patients. However, research models of sepsis
concentrations do not. One can speculate that differ-
and hemorrhagic shock suggest the presence of RAI in
ences in plasma cortisol concentrations may have been
animals.21,22 There are also a small number of clinical
observed if plasma samples had been obtained at later
studies that suggest RAI or RAI-like syndromes in
times. The higher ACTH concentrations in MDR1 wild-
certain populations of veterinary patients.f,g,h
type dogs may have resulted in greater plasma cortisol
Interestingly, herding breed dogs such as Collies and
concentrations after the last sample time point (i.e., 8
Shelties appear to have a reputation for not handling
hours after dexamethasone administration). It is also
illness well. For example, 1 oncologist stated in an on-
possible that greater subject numbers would have dem-
line discussion that ‘Shelties are indeed ‘‘wimps’’ and
onstrated a significant difference in plasma cortisol
they tend to join Collies in this category.’c Another on-
concentrations. There was also greater variability for
cologist commented that ‘Collies sometimes do not
plasma cortisol concentrations than for plasma ACTH
participate in their own recovery.’c Indeed, the obser-
concentrations. Whether this difference may have been
vation that led to the study reported here involved
due to variability among the subjects or variability
what appeared to be an RAI-like phenomenon in an
within the cortisol assay as compared with the ACTH
MDR1 mutant Collie that had undergone a prolonged,
assay is not known. However, despite the variability of
but relatively benign, surgical procedure.
plasma cortisol concentrations, the number of subjects
From this study, it appears that P-gp is an important
was adequate to demonstrate significant differences in
component of the HPA axis in dogs. Dogs lacking P-gp
HPA axis regulation for several other parameters tested
(i.e., MDR1 mutant dogs) appear to have a chronically
including resting cortisol concentrations, post-ACTH
suppressed HPA axis compared with normal dogs.
cortisol concentrations, and greater suppression of
Further studies are necessary to determine the rela-
ACTH secretion after dexamethasone administration.
tionship between MDR1 genotype and RAI. However,
Collectively, results of this study provide evidence
it might be prudent for veterinarians to consider testing
that the HPA axis in MDR1 mutant dogs is suppressed
for RAI (ACTH stimulation) in critically ill canine pa-
as compared with wildtype dogs. Similar to abcb1ab
tients of the herding breed group, since the MDR1 mu-
( À / À ) double knockout mice, MDR1 mutant dogs
tation has been identified in Collies, Shelties, Old
lack P-gp function. Consequently, there is increased
English Sheepdogs, Australian Shepherds, and oth-
penetration of corticosteroids into the central nervous
ers.23,24 If these dogs have clinical signs consistent with
system, resulting in more pronounced negative feed-
RAI, then ‘physiologic’ doses of corticosteroids may be
back inhibition of stress hormone secretion. While this
considered while awaiting test results. Treatment can be
excessive negative feedback inhibition in MDR1 mutant
gradually discontinued in those dogs that appear to
dogs does not appear to interfere with basal physiologic
respond to treatment and have normal results in re-
function in these dogs, it is reasonable to speculate that
sponse to ACTH stimulation and can be continued in
HPA axis function may be inadequate in situations such
those dogs that fail to respond appropriately. Because
& Veterinary Emergency and Critical Care Society 2006, doi: 10.1111/j.1476-4431.2006.00196.x
MDR1 genotyping is commercially available,a many
7. Seelig A. A general pattern for substrate recognition by P-glyco-
protein. Eur J Biochem 1998; 251:252–261.
owners of Collies and other herding breeds know their
8. Schinkel AH, Smit JJ, van Tellingen O, et al. Disruption of the
dog’s genotype. It is important to obtain this informa-
mouse MDR1a P-glycoprotein gene leads to a deficiency in the
tion and consider the implications of the MDR1 mutant
blood–brain barrier and to increased sensitivity to drugs. Cell1994; 20(77):491–502.
genotype when treating these patients.
9. Mealey KL, Bentjen SA, Gay JM, et al. Ivermectin sensitivity in
collies is associated with a deletion mutation of the MDR1 gene. Pharmacogenetics 2001; 11:727–733.
10. Jonker JW, Wagenaar E, van Deemter L, et al. Role of blood–brain
barrier P-glycoprotein in limiting brain accumulation and sedative
This study was funded by a grant from the Collie
side-effects of asimadoline, a peripherally acting analgaesic drug.
11. Sartor LL, Bentjen SA, Trepanier L, et al. Loperamide toxicity in a
collie with the MDR1 mutation associated with ivermectin sensi-tivity. J Vet Intern Med 2004; 18:117–118.
12. Uhr M, Holsboer F, Muller MB. Penetration of endogenous steroid
hormones corticosterone, cortisol, aldosterone and progesterone
Veterinary Information Network; www.vin.com/members/search.
into the brain is enhanced in mice deficient for both MDR1a and
Veterinary Clinical Pharmacology Laboratory, College of Veterinary
MDR1b P-glycoproteins. J Neuroendocrinol 2002; 14:753–759.
Medicine, Washington State University, Pullman, WA; www.vetmed.
13. Muller MB, Keck ME, Binder EB, et al. ABCB1 (MDR1)-type P-
glycoproteins at the blood–brain barrier modulate the activity of
Endocrine Diagnostic Service, Department of Anatomy, Physiology and
the hypothalamic–pituitary–adrenocortical system: implications
Pharmacology, College of Veterinary Medicine, Auburn University, AL.
Excel 2002, Microsoft Corporation, Redmond, WA.
for affective disorder. Neuropsychopharmacology 2003; 28:1991–
NCSS Number Cruncher Statistical Systems, Kaysville, UT; NCSS.com.
Prittie JE, Barton LJ, Peterson ME, et al. Hypothalmo-pituitary–adrenal
14. Beishuizen A, Thijs LG. Relative adrenal failure in intensive care:
(HPA) axis function in critically ill cats. In: Proceedings of the 9th
an identifiable problem requiring treatment? Best Pract Res Clin
International Veterinary Emergency and Critical Care Symposium, New
Endocrinol Metab 2001; 15:513–531.
Orleans, September 9–13, 2003, p. 771.
15. Marik PE, Zaloga GP. Adrenal insufficiency in the critically ill: a
Farrelly J, Hohenhaus AE, Peterson ME, et al. Evaluation of pituitary–
new look at an old problem. Chest 2002; 122:1784–1796.
adrenal function in cats with lymphoma. In: Proceedings of the 19th
16. Brockus CW, Dillon AR, Kemppainen RJ. Effect of alternate-day
Annual Veterinary Cancer Society Conference, Wood’s Hole, November
activity in dogs. Am J Vet Res 1999; 60:698–702.
Boozer A, Behrend EN, Whitley EM, et al. Hypothalamic–pituitary–
17. Schinkel AH, Wagenaar E, Mol CA, et al. P-glycoprotein in the
adrenal axis function in dogs with neoplasia. J Vet Intern Med 2004;
blood–brain barrier of mice influences the brain penetration and
pharmacological activity of many drugs. J Clin Invest 1996;97:2517–2524.
18. Cordon-Cardo C, O’Brien JP, Casals D, et al. Multidrug-resistance
gene (P-glycoprotein) is expressed by endothelial cells at blood–brain barrier sites. Proc Natl Acad Sci USA 1989; 86:695–698.
1. van Asperen J, Mayer U, van Tellingen O, et al. The functional role
19. Martin LG, Groman RP. Relative adrenal insufficiency in critical
of P-glycoprotein in the blood–brain barrier. J Pharm Sci 1997;
illness. J Vet Emerg Crit Care 2004; 14:149–157.
20. Ligtenberg JJ, Zijlstra JG. The relative adrenal insufficiency
2. Mealey KL, Barhoumi R, Rogers K, et al. Doxorubicin induced
syndrome revisited: which patients will benefit from low-dose
expression of P-glycoprotein in a canine osteosarcoma cell line.
steroids? Curr Opin Crit Care 2004; 10:456–460.
21. Wang P, Ba ZF, Jarrar D, et al. Mechanism of adrenal insufficiency
3. Mealey KL, Bentjen SA, Gay JM, et al. Dexamethasone treatment
following trauma and severe hemorrhage: role of hepatic 11beta-
of a canine, but not human, tumour cell line increases chemore-
hydroxysteroid dehydrogenase. Arch Surg 1999; 134:394–401.
sistance independent of P-glycoprotein and multidrug resistance-
22. Koo DJ, Jackman D, Chaudry IH, et al. Adrenal insufficiency dur-
related protein expression. Vet Comp Oncol 2003; 1:67–75.
ing the late stage of polymicrobial sepsis. Crit Care Med 2001;
4. Page RL, Hughes CS, Huyan S, et al. Modulation of P-glycopro-
tein-mediated doxorubicin resistance in canine cell lines. Antican-
23. Neff MW, Robertson KR, Wong AK, et al. Breed distribution and
history of canine MDR1-1{Delta}, a pharmacogenetic mutation that
5. Mealey KL, Northrup NC, Bentjen SA. Increased toxicity of P-
marks the emergence of breeds from the collie lineage. Proc Natl
glycoprotein-substrate chemotherapeutic agents in a dog with the
Acad Sci USA 2004; 100:11725–11730.
MDR1 deletion mutation associated with ivermectin sensitivity.
24. Nelson OL, Carsten E, Bentjen SA, et al. Ivermectin toxicity in an
J Am Vet Med Assoc 2003; 223:1453–1455, 1434.
Australian Shepherd dog with the MDR1 mutation associated
6. Ford JM, Hait WN. Pharmacologic circumvention of multidrug
with ivermectin sensitivity in Collies. J Vet Intern Med 2003;
resistance. Cytotechnology 1993; 12:171–212.
& Veterinary Emergency and Critical Care Society 2006, doi: 10.1111/j.1476-4431.2006.00196.x
Patient Name : Insurance Information Primary Insurance : Secondary Insurance : Worker’s Comp Insurance * Motor-Vehicle Accident Insurance (circle one) I understand and agree that all services rendered are charged directly to me and that I am personally responsible for payment in the event that my claim for Workers Compensation benefits, Auto Claim, or Insurance is denied. C

 Exogenous substances are not the only substrates for
P-gp. In rodents, endogenous hormones including cor-ticosterone are substrates for P-gp suggesting thatP-gp may have a role in regulating their plasma con-centrations.12 Results from recent studies support this
contention. In rodent studies, P-gp was shown to re-strict access of corticosterone and cortisol to the brain.12Furthermore, abcb1ab ( À / À ) double knockout micethat lack P-gp have a suppressed hypothalamic–pitu-itary–adrenal (HPA) axis compared with wildtype
mice.13 Collectively, these results suggest that P-gpnormally limits the concentration of cortisol and corti-
costerone at the hypothalamus and pituitary bluntingfeedback inhibition of the HPA axis. If this premise is
correct, then higher concentrations of cortisol and cor-ticosterone would be expected to reach the hypo-thalamus and pituitary in animals that lack P-gp re-sulting in greater feedback inhibition of the HPA axis
and low endogenous cortisol levels (Figure 1). Conse-
quently, dogs such as herding breed dogs with theMDR1-1D mutation would be expected to be predis-posed to relative adrenal insufficiency (RAI).
Exogenous substances are not the only substrates for
P-gp. In rodents, endogenous hormones including cor-ticosterone are substrates for P-gp suggesting thatP-gp may have a role in regulating their plasma con-centrations.12 Results from recent studies support this
contention. In rodent studies, P-gp was shown to re-strict access of corticosterone and cortisol to the brain.12Furthermore, abcb1ab ( À / À ) double knockout micethat lack P-gp have a suppressed hypothalamic–pitu-itary–adrenal (HPA) axis compared with wildtype
mice.13 Collectively, these results suggest that P-gpnormally limits the concentration of cortisol and corti-
costerone at the hypothalamus and pituitary bluntingfeedback inhibition of the HPA axis. If this premise is
correct, then higher concentrations of cortisol and cor-ticosterone would be expected to reach the hypo-thalamus and pituitary in animals that lack P-gp re-sulting in greater feedback inhibition of the HPA axis
and low endogenous cortisol levels (Figure 1). Conse-
quently, dogs such as herding breed dogs with theMDR1-1D mutation would be expected to be predis-posed to relative adrenal insufficiency (RAI).
 with a protease inhibitor, 100 mL aprotinin, added. Plas-ma was harvested by centrifugation at 4 1C. Plasma wastransferred to prechilled cryovials and frozen at
À 80 1C until analyzed. Cortisol and ACTH concentra-
tions were determined by a commercial laboratoryc us-ing methodology validated for the dog as previouslydescribed.16 A washout period of at least 2 weeks wasallowed between each study.
with a protease inhibitor, 100 mL aprotinin, added. Plas-ma was harvested by centrifugation at 4 1C. Plasma wastransferred to prechilled cryovials and frozen at
À 80 1C until analyzed. Cortisol and ACTH concentra-
tions were determined by a commercial laboratoryc us-ing methodology validated for the dog as previouslydescribed.16 A washout period of at least 2 weeks wasallowed between each study.