Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Tecnoamb.com.br
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Aug. 2004, p. 4720–4726
0099-2240/04/$08.00ϩ0 DOI: 10.1128/AEM.70.8.4720–4726.2004
Copyright 2004, American Society for Microbiology. All Rights Reserved.
Benzene-Toluene-Ethylbenzene-Xylene–Ethanol
Marcio L. B. Da Silva and Pedro J. J. Alvarez*
Department of Civil and Environmental Engineering, University of Iowa, Iowa City, Iowa 52242
Received 5 March 2004/Accepted 27 April 2004
Methanogenic flowthrough aquifer columns were used to investigate the potential of bioaugmentation to enhance anaerobic benzene-toluene-ethylbenzene-xylene (BTEX) degradation in groundwater contaminated with ethanol-blended gasoline. Two different methanogenic consortia (enriched with benzene or toluene and o-xylene) were used as inocula. Toluene was the only hydrocarbon degraded within 3 years in columns that were not bioaugmented, although anaerobic toluene degradation was observed after only 2 years of acclimation. Significant benzene biodegradation (up to 88%) was observed only in a column bioaugmented with the benzene-enriched methanogenic consortium, and this removal efficiency was sustained for 1 year with no significant decrease in permeability due to bioaugmentation. Benzene removal was hindered by the presence of toluene, which is a more labile substrate under anaerobic conditions. Real-time quantitative PCR analysis showed that the highest numbers of bssA gene copies (coding for benzylsuccinate synthase) occurred in aquifer samples exhibiting the highest rate of toluene degradation, which suggests that this gene could be a useful biomarker for environmental forensic analysis of anaerobic toluene bioremediation potential. bssA continued to be detected in the columns 1 year after column feeding ceased, indicating the robustness of the added catabolic potential. Overall, these results suggest that anaerobic bioaugmentation might enhance the natural attenuation of BTEX in groundwater contaminated with ethanol-blended gasoline, although field trials would be needed to demonstrate its feasibility. This approach may be especially attractive for removing benzene, which is the most toxic and commonly the most persistent BTEX compound under anaerobic conditions.
The widespread contamination of surface and groundwater
ethanol, typically exceeding 4,000 mg literϪ1 near the source
resources by the gasoline oxygenate methyl tert-butyl ether
(41, 42), could make the delivery of sufficient oxygen (which
(MTBE) is leading to its phaseout. Ethanol, a likely candidate
has a relatively low solubility) a technically difficult if not pro-
to substitute MTBE, is increasingly being used as a gasoline
hibitively expensive task. Furthermore, oxygen addition could
additive to meet renewable fuel and Clean Air Act require-
lead to clogging problems due to the precipitation of metal
ments (48). Therefore, the presence of ethanol in groundwater
oxides (30, 61), particularly when the dissolved Fe(II) concen-
contaminated with the gasoline constituents benzene, toluene,
tration exceeds 20 mg literϪ1 (55). Thus, reductive dissolution
ethylbenzene, and xylenes (BTEX) is expected to increase in
of iron (possibly exacerbated by the anaerobic degradation of
ethanol) would increase the difficulty to distribute oxygen and
Previous studies have shown that the preferential degrada-
nutrients required to stimulate aerobic BTEX degradation.
tion of ethanol and the accelerated depletion of nutrients and
Enhanced anaerobic BTEX biodegradation has been re-
electron acceptors that would otherwise be available for hy-
ported following the addition of nitrate (5, 17, 19), chelated
drocarbon biodegradation are likely to hinder BTEX removal
Fe(III) (37, 39), and sulfate (3, 38). Nevertheless, anaerobic
(20, 50). These conditions could contribute to longer BTEX
biostimulation may not be sufficient to ensure BTEX degrada-
plumes, increasing the probability that a potential downgradi-
tion if the aquifer material does not contain specific degraders
ent receptor will be exposed (47, 51). Thus, ethanol could deter
in sufficient numbers to exert measurable degradation rates. In
the acceptability of the natural attenuation for controlling the
such cases, the addition of anaerobic microorganisms with the
migration of BTEX plumes at some sites, leaving a margin for
desired catabolic capacity directly into the contaminated zone
alternative bioremediation approaches to solve the problem.
should be evaluated for its ability to enhance the natural at-
Aerobic bioremediation typically exhibits broader catabolic
tenuation of BTEX and ethanol mixtures.
range and faster BTEX degradation kinetics than anaerobic
The benefits of bioaugmentation have been demonstrated in
systems (20, 50). However, aerobic processes are not univer-
field tests for a wide variety of contaminants, including MTBE
sally applicable, and anaerobic strategies might be more ap-
(52) and chlorinated solvents (23, 24, 26, 40). Similarly, ben-
propriate to treat some ethanol-blended gasoline releases.
zene degradation in a sulfate-reducing zone of a petroleum-
Specifically, the high biochemical oxygen demand exerted by
contaminated aquifer was observed only after the inoculationof a benzene-oxidizing, sulfate-reducing enrichment fromaquatic sediments (59). These studies, however, did not deal
* Corresponding author. Present address: Department of Civil and
with the high electron acceptor demand that is exerted during
Environmental Engineering, Rice University, MS 317, Houston, TX
77251-1892. Phone: (713) 348-5903. Fax: (713) 348-5203. E-mail:
ethanol degradation, which is likely to drive the system rapidly
to methanogenic conditions (22). To date, the ubiquity of
DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
TABLE 1. Primers and probe sequences used in RTQ-PCR
5ЈACGACGGYGGCATTTCTC3Ј 5ЈGCATGATSGGYACCGACA3Ј
FAM-5ЈCTTCTGGTTCTTCTGCACCTTGGACACC3Ј-TAMRA
5ЈCGGTGAATACGTTCYCGG3Ј 5ЈGGWTACCTTGTTACGACTT3Ј
5ЈCGGTGAATACGTCCCTGC3Ј 5ЈAAGGAGGTGATCCTGCCGCA3Ј FAM-5ЈCTTGTACACACCGCCCGTC3Ј-BHQ-1
Phage ()a 5ЈACGCCACGCGGGATG3Ј
5ЈAGAGACACGAAACGCCGTTC3Ј TET-5ЈACCTGTGGCATTTGTGCTGCCG3Ј-TAMRA
a The forward and reverse primer as well as the probe were designed by Beller et al. (7). b The forward primer BACT1369F, reverse primer PROK1492R, and probe TM1389F were developed by Suzuki et al. (53). c The forward primer ARCHMIX1369F (ARCH1-1369F and ARCH2-1369F), reverse primer PROK1541R, and probe TM1389F were developed by Suzuki et al. d The reporter dye used was FAM (6-carboxyfluorescein) or TET (tetrachloro-6-carboxyfluorescein), and the quencher dye was either TAMRA (6-carboxy-
tetramethyl rhodamine) or BlackHole Quencher-1.
methanogenic consortia capable of degrading benzene has not
Previous molecular characterization of the toluene- and o-xylene-enriched
been established (36), and no previous studies have addressed
consortium showed the presence of two archaea (a Methanosaeta sp. and a
how to enhance BTEX biodegradation under methanogenic
Methanospirillum sp.), one sulfate-reducing bacterium (a Desulfotomaculum sp.),and one bacterium not related to any known genus (27). The Methanosaeta sp. is
an obligatory aceticlastic methanogen, oxidizing the carboxylic group of acetate
This paper addresses the potential of bioaugmentation to
to CO2 and reducing the methyl group to methane. The Methanospirillum sp.
enhance the anaerobic degradation of BTEX-ethanol mixtures
uses formate and hydrogen as electron donors to reduce CO2 to methane. The
under methanogenic conditions that are characteristic of sites
Desulfotomaculum sp. is capable of growing acetogenically on ethanol, propi-
contaminated by ethanol-blended gasoline. An emphasis was
onate, butyrate, benzoate, and other metabolites. The limited substrate range ofmethanogens and the observed inhibitory effect of sulfate on toluene degrada-
placed on quantifying the presence of bssA and determining
tion implied that the unknown bacterium might be responsible for initiating
whether its concentration was correlated to toluene degrada-
toluene degradation. BSSA, which is the only enzyme known to initiate anaer-
tion activity. This gene codes for benzylsuccinate synthase
obic toluene degradation, was previously detected in this consortium (6).
(BSSA), a ubiquitous enzyme that initiates the anaerobic deg-
Molecular analysis of the methanogenic, benzene-enriched consortium re-
radation of toluene and xylenes by catalyzing the addition of
vealed four archaeal clones (that grouped with acetoclastic and hydrogenotro-phic methanogens) and two predominant bacterial clones (54). One of these
fumarate to the methyl group (1, 6, 9, 10, 12, 33, 49). The bssA
bacteria grouped with the Desulfosporosinus sp. (which typically utilizes lactate,
gene is expressed in phototrophic (62), denitrifying (8), sulfate-
pyruvate, ethanol, or certain fatty acids as electron donors, reducing sulfate to
reducing (9), and iron-reducing (31) bacteria. However, no
hydrogen sulfide), and the second bacterium grouped with Desulfobacterium
previous studies have determined whether higher numbers of
aniline (which can utilize aniline and phenol as substrates). Based on the phy-
bssA gene copies correspond to higher toluene degradation
logenetic association of D. aniline with a clone found in a sulfate-reducing culturecapable of benzene degradation (45), and recognizing that sulfate-reducing bac-
rates under methanogenic conditions, which is important for
teria can grow fermentatively in the absence of sulfate as an electron acceptor
the evaluation of the usefulness of this gene as a biomarker for
(16), Ulrich and Edwards (54) postulated that the latter microorganism initiates
environmental forensic analysis of anaerobic bioremediation
benzene degradation in this consortium.
To date, no toluene-, o-xylene-, or benzene-degrading organism has been
isolated from either of these two consortia. Column inoculation. Two columns exposed to BTEX and ethanol, which had MATERIALS AND METHODS
not exhibited BTEX degradation within 1 year were inoculated with methano-
Aquifer columns. Flowthrough aquifer columns (120-cm length, 5-cm diame-
genic consortia. One column was bioagumented with the toluene- and o-xylene-
ter) equipped with eight sampling ports (at 2.5, 7.6, 14, 20, 40, 60, 80, and 100 cm
enriched consortium, and another was inoculated with the benzene-enriched
from the inlet) were used to investigate the effect of ethanol on BTEX natural
culture. About 40 ml of the cell stock solution (ϳ107 cells mlϪ1) was injected into
attenuation (22) and their enhanced biodegradation through anaerobic bioaug-
a port vial located 20 cm from the column inlet. This location was selected to
mentation. The columns were packed with aquifer material collected from the
determine whether specific BTEX degraders would migrate towards the col-
Northwest Terminal in Tigard, Oregon, and were fed continuously over 3 years
umn’s inlet, where BTEX and ethanol concentrations are higher, or away from
in an up-flow mode with ethanol (ϳ1,000 mg literϪ1) and BTEX (total of 13 to
the source, where ethanol no longer remained (22). The third column, which was
26 mg literϪ1) dissolved in a carbonate-buffered mineral medium. The substrates
not bioaugmented, served as a control to discern the benefits of bioaugmenta-
were fed by using a syringe pump (Harvard Apparatus model 22) and the
tion. The fourth column was poisoned with 0.01% of Kathon CG/ICP biocide
carbonate medium was fed by using a peristaltic pump (Masterflex model 7519-
(5-chloro-2-methyl-3(2H)-isothiazolone and 2-methyl-3(2H)-isothiazolone solu-
15). The ratio of peristaltic pump to syringe pump was set at 20:1. The mineral
tion; Sigma-Aldrich) to distinguish biodegradation from abiotic losses (i.e., vol-
medium composition (in milligrams per liter) was as follows: NOϪ
atilization). All columns were operated in the dark and at an average tempera-
(4.0); CaCO3 (1,000); NH4 (5.5); Mg2ϩ (1.5); PO4 (0.06); and Ni(II),
Cu(II), Zn(II), Co(II), and Mo(IV) (0.002 each). The medium was constantly
Soil DNA extraction. Column aquifer material was collected for DNA analy-
ses. About 5 to 10 g of soil was taken from the port vials located 2.5, 14, 20, and
2-CO2 (95:5, vol/vol) gas to remove dissolved oxygen. The hydrau-
lic characteristics of the columns were estimated by fitting bromide tracer data to
60 cm from the columns’ inlet. Soil samples were dried overnight at room
the one-dimensional advection-dispersion equation as described previously (22),
temperature (22°C), and 0.5 g of the dry soil was then transferred into a lysing
and the hydraulic parameters (with values in parentheses) are as follows: flow
matrix tube for DNA extraction by using a FastDNA SPIN kit according to the
(6.7 to 7.5 ml hϪ1), effective porosity (0.37), dispersion (5 cm2 hϪ1), and seepage
manufacturer’s protocols. A bead-beating device (MINI Beadbeater) was uti-
lized for soil lysis. A 50-l soil DNA sample was collected in a 1.5-ml Eppendorf
Cultures for bioaugmentation. Two methanogenic BTEX-enriched consortia
vial and stored in a freezer (ScienTemp) at Ϫ44°C. RTQ-PCR. The total numbers of bacteria, archaea, and bssA gene copies were
2 and CH4 (27, 54) were utilized as inocula.
The toluene- and o-xylene-enriched consortium was obtained from a site con-
estimated by using real-time quantitative PCR (RTQ-PCR) analysis with primers
taminated with creosote and maintained in the laboratory for 10 years (27). The
and probes (Integrated DNA Technologies, Inc.) described in Table 1. Bacte-
benzene-enriched consortium was obtained from an oil refinery site (54). This
riophage (500 bp) was used as an internal standard for the determination of
consortium was derived from sulfate-reducing microcosms that became metha-
DNA efficiency recovery. When the recovery was lower than 100%, gene copy
nogenic after 1.5 years and was maintained in the lab for six subsequent years.
numbers were normalized to the fraction recovered. No correction was made
Bromide, acetate, nitrate, and sulfate were analyzed in an Alcott 728 auto
sampler equipped with a Gilson 307 pump and a Dionex IonPac AS4A 4-mmcolumn. A Dionex ASRS-I 4-mm anion self-regenerating suppressor and aDionex conductivity detector ion chromatograph were connected to an interface(HP35900E) and a conductivity detector (Ionpac AS4A column). The eluentsolution consisted of 250 mg of sodium bicarbonate literϪ1 plus 933 mg of so-dium carbonate literϪ1 in deionized water. The regenerant solution consisted ofdeionized water with 2.68 ml of concentrated sulfuric acid (18 M) added per liter. The pH of the samples was measured by using a Fisher Scientific AB15 pHmeter. RESULTS AND DISCUSSION Column operation history. Flowthrough aquifer columns
were used to simulate the bioattenuation of BTEX and ethanol
mixtures over 3 years. The removal of BTEX or ethanol was
FIG. 1. Example of calibration curve used for RTQ-PCR. Data
attributable to biodegradation, since negligible abiotic losses
show the standardization of the bssA gene by using genomic DNA of
for BTEX and ethanol (Ͻ8%) were observed in the poisoned
T. aromatica strain T1 (ATCC 700265D). The primers and probes used
control column (data not shown). Methane concentrations
measured in these columns (18 to 23 mg literϪ1) were close to
the solubility level (24 mg literϪ1 at 1 atm and 20°C) (Fig. 2B
and 3B), confirming that methanogenic conditions prevailed.
when DNA recoveries exceeded 100%. Recoveries for the internal standard in
Toluene was the only hydrocarbon that was biodegraded in
the column bioaugmented with benzene-enriched consortium were 115% (at 2.5
the column that was not bioaugmented. However, its degrada-
cm), 14% (at 14 cm), and 4% (at 60 cm). Recoveries for the internal standard inthe column bioaugmented with toluene- and o-xylene-enriched consortium were
tion occurred after only 2 years of acclimation (ϳ137 pore
148% (at 2.5 cm), 4% (at 14 cm), 6% (at 20 cm), and 15% (at 60 cm). Such wide
volumes exchanged) (data not shown). On the other hand,
variations in DNA recoveries (e.g., 0.6 to 126%) are commonly reported (44, 63)
benzene degradation was not observed within 3 years. The
and are probably due to the binding of sample impurities (e.g., humic acids) that
overall persistence of BTEX compounds was attributed to the
interfere with the activity of Taq polymerase during PCR analysis (46).
The PCR mixture contained a 0.9 M concentration of each primer (a 0.45
development of strongly anaerobic (methanogenic) conditions
M concentration of each forward primer for archaea), a 0.25 M concentration
as result of the high electron acceptor demand exerted during
of the probe, 1ϫ TaqMan Universal PCR Master mix (Applied Biosystems), 2.5
ethanol degradation, which depleted the influent nitrate and
l of undiluted DNA, and nuclease-free sterile water (AMRESO-E476) to a final
sulfate. The low oxidation-reduction found in these columns
volume of 25 l. The RTQ-PCR was conducted with an ABI PRISM 7000
sequence detection system (Applied Biosystems) with the following temperature
h ϭ Ϫ300 mV) (22) represents decreased thermodynamic
conditions: 50°C for 2 min, followed by 95°C for 10 min and 40 cycles at 95°C for
feasibility for BTEX bio-oxidation. The recalcitrance of ben-
15 s, and 60°C for 1 min. The initial concentration of DNA in the standards was
zene, which is the most toxic and the least frequently degraded
measured by using a Beckman DU-600 fluorometer (Amersham Pharmacia,
of the BTEX compounds under anaerobic conditions (2, 29),
motivated us to investigate whether anaerobic bioaugmenta-
The number of bssA gene copies in each sample was estimated based on the
tion could enhance its natural attenuation in the presence of
Effect of bioaugmentation. Major concerns about the use of
bioaugmentation include the survival of the added microor-
ganisms (34, 43) and/or low bacterial transport through the
aquifer material, which acts as a filter (24). However, bioaug-
This equation is based on the following assumptions: (i) the bssA primer and
mentation has a greater probability of success when the added
probe set designed on different denitrifying bacteria (i.e., Azoarcus sp. strain T,
microorganisms fill a metabolic niche that is not being ex-
Thauera aromatica strains T1 and K172, and Azoarcus tolulyticus strain Tol-4)
ploited by the indigenous microfloras (58). Since aerobic
was representative of all other bacteria containing bssA; (ii) the approximate sizeof the strain T1 genome used as the standard in the calibration curves was 4.6
BTEX-degrading organisms are ubiquitous (28), bioaugmen-
Mbp (and there are approximately 9.12576 ϫ 1014 bp g of DNAϪ1), which is
tation is generally considered to be unnecessary to enhance
equivalent to the size of the Escherichia coli genome (13); and (iii) there is one
aerobic BTEX bioremediation. Nonetheless, the common per-
copy of bssA per genome. These assumptions were also used to quantify bacteria
sistence of benzene in strongly anaerobic environments (exac-
and archaea but not bacteriophage , because, in this case, the solutions con-
erbated by the presence of ethanol) suggests that indigenous
tained DNA fragments of identical length as those used in the standards. Figure1 shows an example of a calibration curve prepared for bssA. Similar calibration
microorganisms do not readily exploit benzene degradation as
curves were also prepared for bacteria (T. aromatica strain T1 [102 to 108 copies;
an ecological niche under methanogenic conditions. There-
r2 ϭ 0.988]), archaea (Methanococcus maripaludis [103 to 108 copies; r2 ϭ
fore, bioaugmentation with competent methanogenic consortia
0.990]), and bacteriophage (101 to 107 copies; r2 ϭ 0.986).
may be justified in such cases to shorten long acclimation
Analytical procedures. Samples for BTEX, ethanol, and methane analyses
were collected with a 1-ml gas-tight syringe directly from the columns’ port vials
periods and enhance degradation rates.
and injected into 5-ml gas chromatography (GC) vials previously capped with a
Figure 2A and B shows BTEX and ethanol concentration
20-mm Teflon-coated septum and aluminum crimps. The vials were placed in a
profiles, respectively, in the column bioaugmented with the
water bath (Buchi 461) at 85°F for 30 min. A 1-ml headspace sample was then
toluene- and o-xylene-enriched methanogenic consortium. No
injected directly into a Hewlett-Packard model 5890 series II GC equipped with
BTEX removal had been observed prior to bioaugmentation.
a DB-wax column (30 m, 0.53-mm diameter; J & W Scientific) and a flameionization detector. GC operational temperatures were set at 175°C for the
The removal rate of toluene and o-xylene occurred within 30
injector, 250°C for the detector, and 150°C for the oven.
days after bioaugmentation and was most pronounced near the
DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
FIG. 2. Concentration profiles in the column bioaugmented with the toluene- and o-xylene-enriched methanogenic consortium, taken 1 year
after inoculation. Concentrations of BTEX (A) and of ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along
the length of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.
inoculation port (located 20 cm from the inlet). BSSA, which is
toluene concentration was decreased from 6.8 to 1.0 mg literϪ1
the enzyme responsible for initiating toluene degradation by
(Fig. 4). A subsequent increase in the influent toluene concen-
the added consortium (6), was probably responsible for the
tration to 2 mg literϪ1 resulted in a decrease in removal effi-
degradation of o-xylene during bacterial growth on toluene,
ciency to about 75% (data not shown). These results suggest
possibly cometabolically as discussed previously (11). Toluene-
that toluene inhibited anaerobic benzene degradation. Com-
dependent degradation of o-xylene appears to be a common
petitive inhibition is unlikely because anaerobic benzene deg-
occurrence in anaerobic systems (2). Interestingly, m- and p-
radation is not mediated by BSSA, which attacks methyl
xylene degradation was not observed. Whereas the initial bio-
groups that benzene lacks. Rather, anaerobic benzene degra-
transformations of m-, p-, and o-xylene have been reported to
dation has been reported to occur through oxidation of the
be analogous to the biotransformation of toluene by BSSA (8,
aromatic ring to form phenol (57, 60) and, subsequently, ben-
11, 12, 33), differences in substrate specificity among BSSA
zoate and a variety of aliphatic acids (29). It is unknown if
enzymes expressed in different organisms are not well under-
toluene was degraded by the same organisms that degraded
stood. For example, all of the m-xylene-anaerobic degrading
benzene and, if so, whether toluene acted as a noncompetitive
bacteria isolated thus far can degrade toluene, but it is unclear
inhibitor, perhaps by being utilized preferentially or contribut-
why the reverse is not always observed (29). Apparently, the
ing to metabolic flux dilution (35). o-Xylene degradation was
BSSA expressed in this consortium is selective towards toluene
also observed in this column 90 days after inoculation, and its
removal increased over time (up to 99%). Overall, the high
Benzene and ethylbenzene were not degraded in this column
BTEX removal efficiency after bioaugmentation was sustain-
(Fig. 2A). The persistence of these compounds corroborates
previous studies showing that this consortium is unable to
Acetate, a by-product of ethanol anaerobic degradation, was
degrade them (25). Benzene (which lacks a methyl group) and
observed in all columns (580 to 650 mg literϪ1) near the inlet,
ethylbenzene (which is transformed by the dehydrogenation of
where the highest ethanol degradation activity occurred (Fig.
the ethyl group) are degraded by different anaerobic pathways
2B and 3B). Acetate accumulation resulted in the reduction of
pH from 8.3 to 6.3 (data not shown), even though the medium
Significant benzene degradation (Ͼ49%) was found only in
was well buffered with calcium carbonate (ϳ1 g literϪ1). Ed-
the column bioaugmented with benzene-enriched microorgan-
wards and Grbic´-Galic´ (25) found that acetate, which is also a
isms (Fig. 3A). Interestingly, benzene removal efficiency in-
by-product of toluene degradation by the toluene- and o-
creased from 49% Ϯ 14% to 88% Ϯ 7% after the influent
xylene-enriched methanogenic consortium, inhibited toluene
FIG. 3. Concentration profiles of the column bioaugmented with the benzene-enriched methanogenic consortium, taken 1 year after inocu-
lation. Concentrations of BTEX (A) and ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along the length
of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.
degradation. Characterizing and elucidating the inhibitory
archaea (e.g., methanogens) (ϳ1010 cells g of soilϪ1) were
mechanisms of acetate on BTEX degradation was beyond the
higher near the column inlet and decreased along the column
scope of this study. However, it did not escape our attention
length as the substrate concentration decreased (Fig. 2 and 3).
that high benzene, toluene, and o-xylene removal rates were
Total bacterial concentrations were also estimated indepen-
continuously observed near the inoculation port where neither
dently for these samples by using phospholipid fatty acid anal-
ethanol nor acetate was present (Fig. 2B and 3B).
The increase in biomass concentration due to bioaugmenta-
tion could potentially decrease soil permeability and affect
nutrient and substrate transport to the growing cells (18, 21,
56). However, no clogging was observed within 1 year of inoc-
ulation, suggesting that bioaugmentation (with 40 ml at ϳ1010
cells literϪ1) had a negligible effect on column permeability.
For example, a microbial concentration (x) on the order of 1010
cells g of soilϪ1, which was the highest measured value (Fig. 2C
and 3C), would decrease soil porosity by only about 0.2%,
assuming a dry cell weight (DCW) of 1.33 ϫ 10Ϫ13 g (14), a soil
bulk density (bulk) of 1,600 g literϪ1, and a biomass density
(cell) of 1,100 g literϪ1 (15), i.e., the pore volume fraction
occupied by the microorganisms is calculated as (xϫ DCW ϫ
Molecular analysis. Soil DNA analysis was performed in the
aquifer material prior to inoculation and after 1 year of exper-
imentation. RTQ-PCR was used to estimate the total number
FIG. 4. Effect of toluene on anaerobic benzene degradation. Ben-
of archaea, bacteria, and specific toluene degraders harboring
zene removal increased from 49 to 88% after the influent toluene
concentration was decreased from 7 to 1 mg literϪ1. Symbols: f,
influent toluene of 6.75 Ϯ 1.6 mg literϪ1; ᮀ, influent toluene of 1.1 Ϯ
The total numbers of bacteria (ϳ108 cells g of soilϪ1) and
DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
bioaugmentation is unlikely to be universally applicable, and
pilot studies should be conducted to identify potential critical
limitations associated with scale-up issues, including the re-
quired inoculum size and cost, the need for pH control, and
performance at lower temperatures in the presence of poten-
ACKNOWLEDGMENTS
We thank Elizabeth Edwards and Ania Ulrich (University of To-
ronto, Toronto, Canada) for donating the bacterial consortia utilized
in this work, Tim Buschek for donating the aquifer material, Harry
Beller and Stacy Kane from Lawrence Livermore National Laboratory
for their comments and suggestions regarding RTQ-PCR, Craig Just
FIG. 5. Persistence of the bssA gene in soil samples from the col-
and Collin Just (University of Iowa) for their analytical assistance, and
umn bioaugmented with toluene- and o-xylene-enriched methanogenic
the DNA Facility (University of Iowa) for its help with PCR analysis.
consortium. Open bars, 1 year after inoculation; filled bars, 1 year after
This work was funded by a grant from the U.S. Environmental
exposure to BTEX and ethanol (EtOH) ceased.
Protection Agency STAR program. M.L.B.D.S. was also supported by
ysis by Microbial Insights, Inc. Its results (5 ϫ 107 to 7 ϫ 107
REFERENCES
cells g of soilϪ1) were in good agreement with our RTQ-PCR
1. Achong, G. R., A. M. Rodriguez, and A. M. Spormann. 2001. Benzylsuccinate
synthase of Azoarcus sp. strain T: cloning, sequencing, transcriptional orga-
Little is known about the diversity of the bssA gene in dif-
nization, and its role in anaerobic toluene and m-xylene mineralization.
ferent bacteria. Therefore, it is unclear whether the bssA prim-
J. Bacteriol. 183:6763–6770.
2. Alvarez, P. J. J., and T. M. Vogel. 1995. Degradation of BTEX and their
er-probe combination used in this study (which was designed
aerobic metabolites by indigenous microorganisms under nitrate reducing
based on the genome sequence of denitrifying bacteria) might
conditions. Water Sci. Technol. 31:15–28.
3. Anderson, R. T., and D. R. Lovley. 2000. Anaerobic bioremediation of
require alterations to enhance hybridization with the bssA se-
benzene under sulfate-reducing conditions in a petroleum-contaminated
quences in the methanogenic consortia present in the columns.
aquifer. Environ. Sci. Technol. 34:2261–2266.
A suboptimal primer-probe sequence could have resulted in an
4. Ball, H. A., H. A. Johnson, M. Reinhard, and A. M. Spormann. 1996. Initial
reactions in anaerobic ethylbenzene oxidation by a denitrifying bacterium,
underestimation of the total number of bssA copies. Neverthe-
strain EB1. J. Bacteriol. 178:5755–5761.
less, the primer-probe set used in this work reproductively
5. Ball, H. A., and M. Reinhard. 1996. Monoaromatic hydrocarbon transfor-
detected bssA (detection limit of Ն10 bssA copies g of soilϪ1)
mation under anaerobic conditions at Seal Beach, California: laboratory
studies. Environ. Toxicol. Chem. 15:114–122.
in both columns, and the highest number of bssA gene copies
6. Beller, H. R., and E. A. Edwards. 2000. Anaerobic toluene activation by
was measured near the location exhibiting the highest toluene
benzylsuccinate synthase in a highly enriched methanogenic culture. Appl.
Environ. Microbiol. 66:5503–5505.
degradation activity (Fig. 2). Therefore, this gene might be
7. Beller, H. R., S. R. Kane, T. C. Legler, and P. J. J. Alvarez. 2002. A real-time
a useful biomarker for environmental forensic analyses to
polymerase chain reaction method for monitoring anaerobic, hydrocarbon-
evaluate anaerobic toluene biodegradation potential. The
degrading bacteria based on a catabolic gene. Environ. Sci. Technol. 36:
detection of bssA near the inlet, albeit at levels 4 orders of
8. Beller, H. R., and A. M. Spormann. 1997. Anaerobic activation of toluene
magnitude lower than that at the point of highest toluene
and o-xylene by addition to fumarate in denitrifying strain T. J. Bacteriol.
degradation activity (Fig. 2C), suggests chemotaxis of anaero-
179:670–676.
9. Beller, H. R., and A. M. Spormann. 1997. Benzylsuccinate formation as a
bic toluene degraders toward areas of higher substrate concen-
means of anaerobic toluene activation by sulfate-reducing strain PRTOL1.
Appl. Environ. Microbiol. 63:3729–3731.
10. Beller, H. R., and A. M. Spormann. 1999. Substrate range of benzylsuccinate
To investigate the persistence of the bssA gene, RTQ-PCR
synthase from Azoarcus sp. strain T. FEMS Microbiol. Lett. 178:147–153.
analysis was performed up to 1 year after column feeding was
11. Beller, H. R., A. M. Spormann, P. K. Sharma, J. R. Cole, and M. Reinhard.
stopped (and thus, exposure to BTEX and ethanol had ceased)
1996. Isolation and characterization of a novel toluene-degrading, sulfate-
reducing bacterium. App. Environ. Microbiol. 62:1188–1196.
(Fig. 5). The results showed that bssA genes were still present
12. Biegert, T., G. Fuchs, and J. Heider. 1996. Evidence that anaerobic oxidation
in the aquifer material (although at lower concentrations, de-
of toluene in the denitrifying bacterium Thauera aromatica is initiated by
creasing by 92% near the inlet and 65% at the inoculation
formation of benzylsuccinate from toluene and fumarate. Eur. J. Biochem. 238:661–668.
port). This detection suggests the robustness of anaerobic bio-
13. Blattner, F. R., G. Plunkett, C. A. Bloch, N. T. Perna, V. Burland, M. Riley,
augmentation, which may offer long-term enhanced biodegra-
J. Collado-Vides, J. D. Glasner, C. K. Rode, G. F. Mayhew, J. Gregor, N. W. Davis, H. A. Kirkpatrick, M. A. Goeden, D. J. Rose, B. Mau, and Y. Shao.
1997. The complete genome sequence of Escherichia coli K-12. Science
In conclusion, the increasing likelihood of encountering eth-
277:1453–1474.
anol as a cocontaminant in groundwater contaminated with
14. Bratbak, G. 1985. Bacterial biovolume and biomass estimations. Appl. En-
viron. Microbiol. 49:1488–1493.
BTEX may require the reconsideration of current remediation
15. Bratbak, G., and I. Dundas. 1984. Bacterial dry-matter content and biomass
practices and the development of novel remediation ap-
estimations. Appl. Environ. Microbiol. 48:755–757.
proaches. This work supports the notion that anaerobic bio-
16. Bryant, M., L. Campbell, C. Reddy, and M. Crabill. 1977. 1977. Growth of Desulfovibrio in lactate or ethanol media low in sulfate in association with
augmentation could be a robust and sustainable approach to
H2-utilizing methanogenic bacteria. Appl. Environ. Microbiol. 33:1162–
accelerate the natural attenuation of BTEX in some aquifers
17. Burland, S. M., and E. A. Edwards. 1999. Anaerobic benzene biodegradation
contaminated with ethanol-blended gasoline. This approach
linked to nitrate reduction. Appl. Environ. Microbiol. 65:529–533.
may be especially attractive for removing benzene, which is the
18. Clement, T. P., B. S. Hooker, and R. S. Skeen. 1996. Macroscopic models for
most toxic and commonly the most persistent BTEX com-
predicting changes in saturated porous media properties caused by microbial
growth. Ground Water 34:934–942.
pound under anaerobic conditions. Nevertheless, anaerobic
19. Coates, J. D., R. Chakraborty, J. G. Lack, S. M. O’Connor, K. A. Cole, K. S. Bender, and L. A. Achenbach. 2001. Anaerobic benzene oxidation coupled to
the impact of ethanol on the persistence of benzene in gasoline-contami-
nitrate reduction in pure culture by two strains of Dechloromonas. Nature
nated groundwater. Water Resour. Res. 38:1003. 411:1039–1043.
43. Munakata-Marr, J., P. L. McCarty, M. S. Shields, M. Reagin, and S. C.
20. Corseuil, H. X., C. S. Hunt, R. C. F. Dos Santos, and P. J. J. Alvarez. 1998. Francesconi. 1996. Enhancement of trichloroethylene degradation in aquifer
The influence of the gasoline oxygenate ethanol on aerobic and anaerobic
microcosms bioaugmented with wild type and genetically altered Burkhold-
BTX biodegradation. Water Res. 32:2065–2072. eria (Pseudomonas) cepacia G4 and PR1. Environ. Sci. Technol. 30:2045–
21. Cunningham, A. B., W. G. Characklis, F. Abedeen, and D. Crawford. 1991.
Influence of biofilm accumulation on porous-media hydrodynamics. Envi-
44. Mygind, T., L. Ostergaard, S. Birkelund, J. S. Lindholt, and G. Chris-
ron. Sci. Technol. 25:1305–1311. tiansen. 2003. Evaluation of five DNA extraction methods for purification of
22. Da Silva, M. L. B., and P. J. J. Alvarez. 2002. Effects of ethanol versus MTBE
DNA from atherosclerotic tissue and estimation of prevalence of Chlamydia
on benzene, toluene, ethylbenzene, and xylene natural attenuation in aquifer
pneumoniae in tissue from a Danish population undergoing vascular repair.
columns. J. Environ. Eng. ASCE 128:862–867.
BMC Microbiol. 3:19.
23. Duba, A. G., K. J. Jackson, M. C. Jovanovich, R. B. Knapp, and R. T. Taylor.
45. Phelps, C. D., L. J. Kerkhof, and L. Y. Young. 1998. Molecular character-
1996. TCE remediation using in situ, resting-state bioaugmentation. Envi-
ization of a sulfate-reducing consortium which mineralizes benzene. FEMS
ron. Sci. Technol. 30:1982–1989.
Microbiol. Ecol. 27:269–279.
24. Dybas, M. J., M. Barcelona, S. Bezborodnikov, S. Davies, L. Forney, H.
46. Porteous, L. A., R. J. Seidler, and L. S. Watrud. 1997. An improved method Heuer, O. Kawka, T. Mayotte, L. Sepulveda-Torres, K. Smalla, M. Sneathen,
for purifying DNA from soil for polymerase chain reaction amplification and
J. Tiedje, T. Voice, D. C. Wiggert, M. E. Witt, and C. S. Criddle. 1998.
molecular ecology applications. Mol. Ecol. 6:787–791.
Pilot-scale evaluation of bioaugmentation for in-situ remediation of a carbon
47. Powers, S. E., C. S. Hunt, S. E. Heermann, H. X. Corseuil, D. Rice, and
tetrachloride contaminated aquifer. Environ. Sci. Technol. 32:3598–3611. P. J. J. Alvarez. 2001. The transport and fate of ethanol and BTEX in
25. Edwards, E. A., and D. Grbic´-Galic´. 1994. Anaerobic degradation of toluene
groundwater contaminated by gasohol. Crit. Rev. Environ. Sci. Technol.
and o-xylene by a methanogenic consortium. Appl. Environ. Microbiol. 60: 31:79–123.
48. Powers, S. E., D. Rice, B. Dooher, and P. J. J. Alvarez. 2001. Will ethanol-
26. Ellis, D. E., E. J. Lutz, J. M. Odom, R. J. Buchanan, C. L. Bartlett, M. D.
blended gasoline affect groundwater quality? Using ethanol instead of
Lee, M. R. Harkness, and K. A. Deweerd. 2000. Bioaugmentation for accel-
MTBE as a gasoline oxygenate could be less harmful to the environment.
erated in situ anaerobic bioremediation. Environ. Sci. Technol. 34:2254–
Environ. Sci. Technol. 35:24A–30A.
49. Rabus, R., and J. Heider. 1998. Initial reactions of anaerobic metabolism of
27. Ficker, M., K. Krastel, S. Orlicky, and E. Edwards. 1999. Molecular char-
alkylbenzenes in denitrifying and sulfate reducing bacteria. Arch. Microbiol.
acterization of a toluene-degrading methanogenic consortium. Appl. Envi-
170:377–384.
ron. Microbiol. 65:5576–5585.
50. Ruiz-Aguilar, G. M. L., J. M. Fernandez-Sanchez, S. R. Kane, D. Kim, and
28. Gibson, D. T., and V. Subramanian. 1984. Microbial degradation of aromatic P. J. J. Alvarez. 2002. Effect of ethanol and methyl-tert-butyl ether on
hydrocarbons, p. 181–252. In D. T. Gibson (ed.), Microbial degradation of
monoaromatic hydrocarbon biodegradation: response variability for differ-
organic compounds. Marcel Dekker, Inc., New York, N.Y.
ent aquifer materials under various electron-accepting conditions. Environ.
29. Heider, J., A. M. Spormann, H. R. Beller, and F. Widdel. 1998. Anaerobic
Toxicol. Chem. 21:2631–2639.
bacterial metabolism of hydrocarbons. FEMS Microbiol. Rev. 22:459–473.
51. Ruiz-Aguilar, G. M. L., K. O’Reilly, and P. J. J. Alvarez. 2002. A comparison
30. Hinchee, R. E., and D. C. Downey. 1988. The role of hydrogen peroxide in
of benzene and toluene plume lengths for sites contaminated with regular vs.
enhanced bioreclamation, p. 931–948. In National Water Well Association
ethanol-amended gasoline. Ground Water Monit. Res. 23:48–53.
(ed.), Proceedings of the NWWA/API Conference on Petroleum Hydrocar-
52. Salanitro, J. P., P. C. Johnson, G. E. Spinnler, P. M. Maner, H. L.
bons and Organic Chemicals in Ground Water: Prevention, Detection and
Wisniewski, and C. Bruce. 2000. Field scale demonstration of enhanced
Restoration. American Petroleum Institute, Houston, Tex.
MTBE bioremediation through aquifer bioaugmentation and oxygenation.
Environ. Sci. Technol. 34:4152–4162. Kane, S. R., H. R. Beller, T. C. Legler, and R. T. Anderson. 2002. Biochem-
ical and genetic evidence of benzylsuccinate synthase in toluene-degrading,
53. Suzuki, M. T., L. T. Taylor, and E. F. DeLong. 2000. Quantitative analysis of
ferric iron-reducing Geobacter metallireducens. Biodegradation
small-subunit rRNA genes in mixed microbial populations via 5 Ј-nuclease
13:149–154.
assays. Appl. Environ. Microbiol. 66:4605–4614.
32. Kniemeyer, O., and J. Heider. 2001. (S)-1-phenylethanol dehydrogenase of
54. Ulrich, A. C., and E. A. Edwards. 2003. Physiological and molecular char- Azoarcus sp. strain EbN1, an enzyme of anaerobic ethylbenzene catabolism.
acterization of anaerobic benzene-degrading mixed cultures. Environ. Mi-
Arch. Microbiol. 129:129–135.
crobiol. 5:92–102.
33. Krieger, C. J., H. R. Beller, M. Reinhard, and A. M. Spormann. 1999. Initial
55. U.S. Environmental Protection Agency. 1985. How to evaluate alternative
reactions in anaerobic oxidation of m-xylene by the denitrifying bacterium
cleanup technologies for underground storage tank sites: a guide for correc-
Azoarcus sp. strain T. J. Bacteriol. 181:6403–6410.
tive action plan reviewers. U.S. Environmental protection agency document
34. Krumme, M. L., R. L. Smith, J. Egestorff, S. M. Thiem, J. M. Tiedje, K. N.
no. 510-B-94–003 and 510-B-95–007. U.S. Environmental Protection
Timmis, and D. F. Dwyer. 1994. Behavior of pollutant-degrading microor-
ganisms in aquifers—predictions for genetically-engineered organisms. En-
56. Vandevivere, P., and P. Baveye. 1992. Relationship between transport of
viron. Sci. Technol. 28:1134–1138.
bacteria and their clogging efficiency in sand columns. Appl. Environ. Mi-
35. Lovanh, N., and P. J. J. Alvarez. 2004. Effect of ethanol, acetate, and phenol
crobiol. 58:2523–2530.
on toluene degradation activity and tod-lux expression in Pseudomonas
57. Vogel, T. M., and D. Grbicgalic. 1986. Incorporation of oxygen from water putida TOD102: evaluation of the metabolic flux dilution model. Biotechnol.
into toluene and benzene during anaerobic fermentative transformation.
Bioeng. 86:801–808.
Appl. Environ. Microbiol. 52:200–202.
36. Lovley, D. R. 2000. Anaerobic benzene degradation. Biodegradation 11:107–
58. Vogel, T. M., and M. V. Water. 2003. Bioaugmentation, p. 952–959. In C. J.
Hurst, R. L. Crawford, G. R. Knudsen, M. J. McInerney, and L. D. Stetzen-
37. Lovley, D. R. 1991. Dissimilatory Fe(III) and Mn(IV) Reduction. Microbiol.
bach (ed.), Manual of environmental microbiology, 2nd ed. ASM Press,
Rev. 55:259–287.
38. Lovley, D. R., J. D. Coates, J. C. Woodward, and E. J. P. Phillips. 1995.
59. Weiner, J. M., and D. R. Lovley. 1998. Anaerobic benzene degradation in
Benzene oxidation coupled to sulfate reduction. Appl. Environ. Microbiol.
petroleum-contaminated aquifer sediments after inoculation with a benzene-
61:953–958.
oxidizing enrichment. Appl. Environ. Microbiol. 64:775–778.
39. Lovley, D. R., J. C. Woodward, and F. H. Chapelle. 1994. Stimulated anoxic
60. Weiner, J. M., and D. R. Lovley. 1998. Rapid benzene degradation in metha-
biodegradation of aromatic-hydrocarbons using Fe(III) ligands. Nature 370:
nogenic sediments from a petroleum-contaminated aquifer. Appl. Environ.
Microbiol. 64:1937–1939.
40. Major, D. W., M. L. McMaster, E. E. Cox, E. A. Edwards, S. M. Dworatzek,
61. Wiesner, M. R., M. C. Grant, and S. R. Hutchins. 1996. Reduced perme- E. R. Hendrickson, M. G. Starr, J. A. Payne, and L. W. Buonamici. 2002.
ability in groundwater remediation systems: Role of mobilized colloids and
Field demonstration of successful bioaugmentation to achieve dechlorina-
injected chemicals. Environ. Sci. Technol. 30:3184–3191.
tion of tetrachloroethene to ethene. Environ. Sci. Technol. 36:5106–5116.
62. Zengler, K., J. Heider, R. Rossello-Mora, and F. Widdel. 1999. Phototrophic
41. Malcolm Pirnie, Inc. 1998. Evaluation of the fate and transport of ethanol in
utilization of toluene under anoxic conditions by a new strain of Blastochloris
the environment. Report prepared for the American Methanol Institute. sulfoviridis. Arch. Microbiol. 172:204–212.
Malcolm Pirnie, Inc., Oakland, Calif.
63. Zhou, J. Z., M. A. Bruns, and J. M. Tiedje. 1996. DNA recovery from soils
42. Molson, J. W., J. F. Barker, E. O. Frind, and M. Schirmer. 2002. Modeling
of diverse composition. Appl. Environ. Microbiol. 62:316–322.
Título do trabalho: A ordem cerebral e as ‘desordens’ do cérebro: uma etnografia da divulgação neurocientífica e da psiquiatria biológica1 Autor: Rogerio Lopes Azize e-mail: rogerioazize@hotmail.com Filiação institucional: Doutorando em Antropologia Social pelo PPGAS/MN/UFRJ GT 19: Natureza, corpo e sentidos Coordenadores: Cynthia Andersen Sarti (UNIFESP), Luiz Fernando Dias Duarte (MN
 Bromide, acetate, nitrate, and sulfate were analyzed in an Alcott 728 auto
sampler equipped with a Gilson 307 pump and a Dionex IonPac AS4A 4-mmcolumn. A Dionex ASRS-I 4-mm anion self-regenerating suppressor and aDionex conductivity detector ion chromatograph were connected to an interface(HP35900E) and a conductivity detector (Ionpac AS4A column). The eluentsolution consisted of 250 mg of sodium bicarbonate literϪ1 plus 933 mg of so-dium carbonate literϪ1 in deionized water. The regenerant solution consisted ofdeionized water with 2.68 ml of concentrated sulfuric acid (18 M) added per liter.
Bromide, acetate, nitrate, and sulfate were analyzed in an Alcott 728 auto
sampler equipped with a Gilson 307 pump and a Dionex IonPac AS4A 4-mmcolumn. A Dionex ASRS-I 4-mm anion self-regenerating suppressor and aDionex conductivity detector ion chromatograph were connected to an interface(HP35900E) and a conductivity detector (Ionpac AS4A column). The eluentsolution consisted of 250 mg of sodium bicarbonate literϪ1 plus 933 mg of so-dium carbonate literϪ1 in deionized water. The regenerant solution consisted ofdeionized water with 2.68 ml of concentrated sulfuric acid (18 M) added per liter.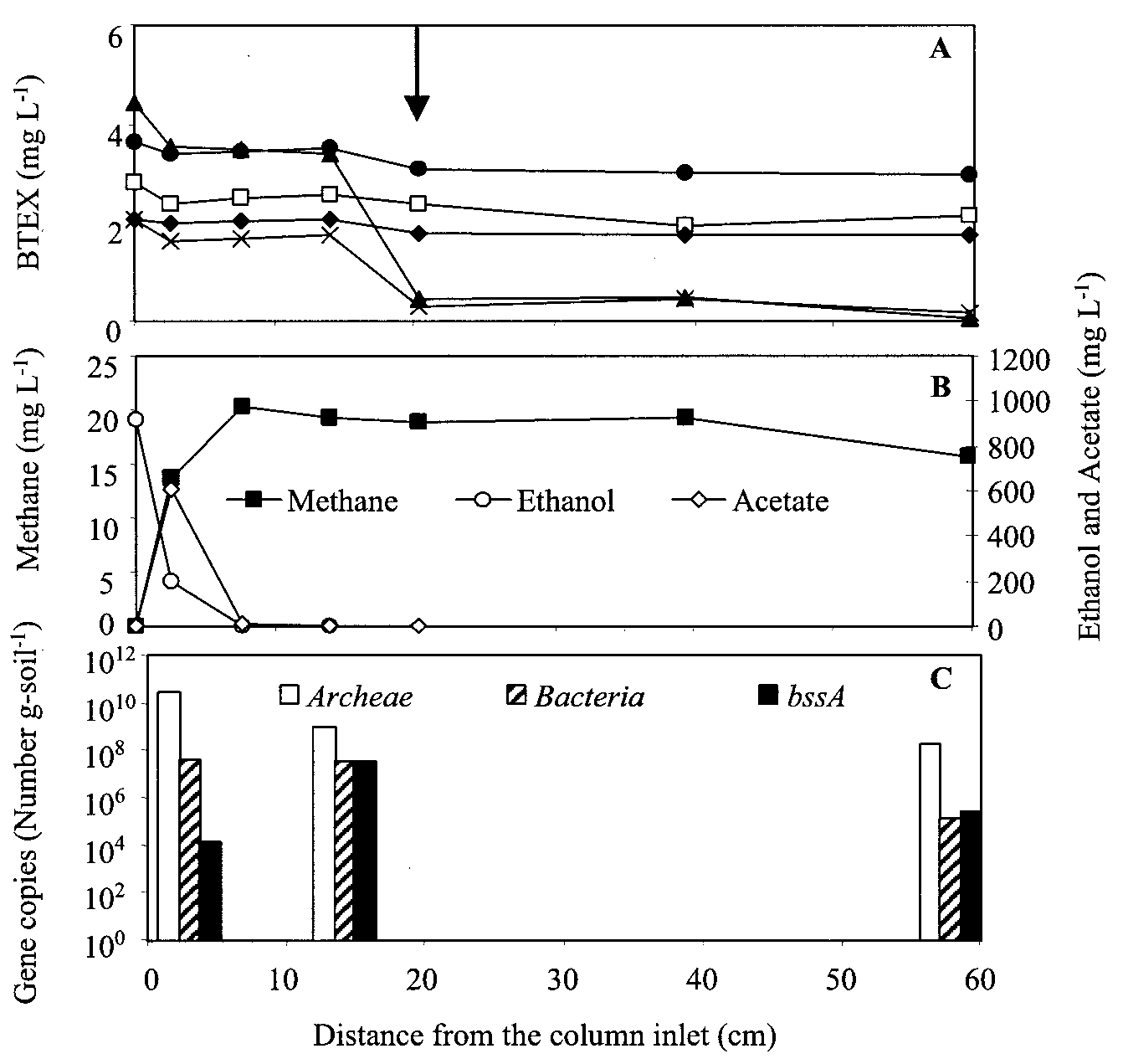 DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
FIG. 2. Concentration profiles in the column bioaugmented with the toluene- and o-xylene-enriched methanogenic consortium, taken 1 year
after inoculation. Concentrations of BTEX (A) and of ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along
the length of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.
DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
FIG. 2. Concentration profiles in the column bioaugmented with the toluene- and o-xylene-enriched methanogenic consortium, taken 1 year
after inoculation. Concentrations of BTEX (A) and of ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along
the length of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.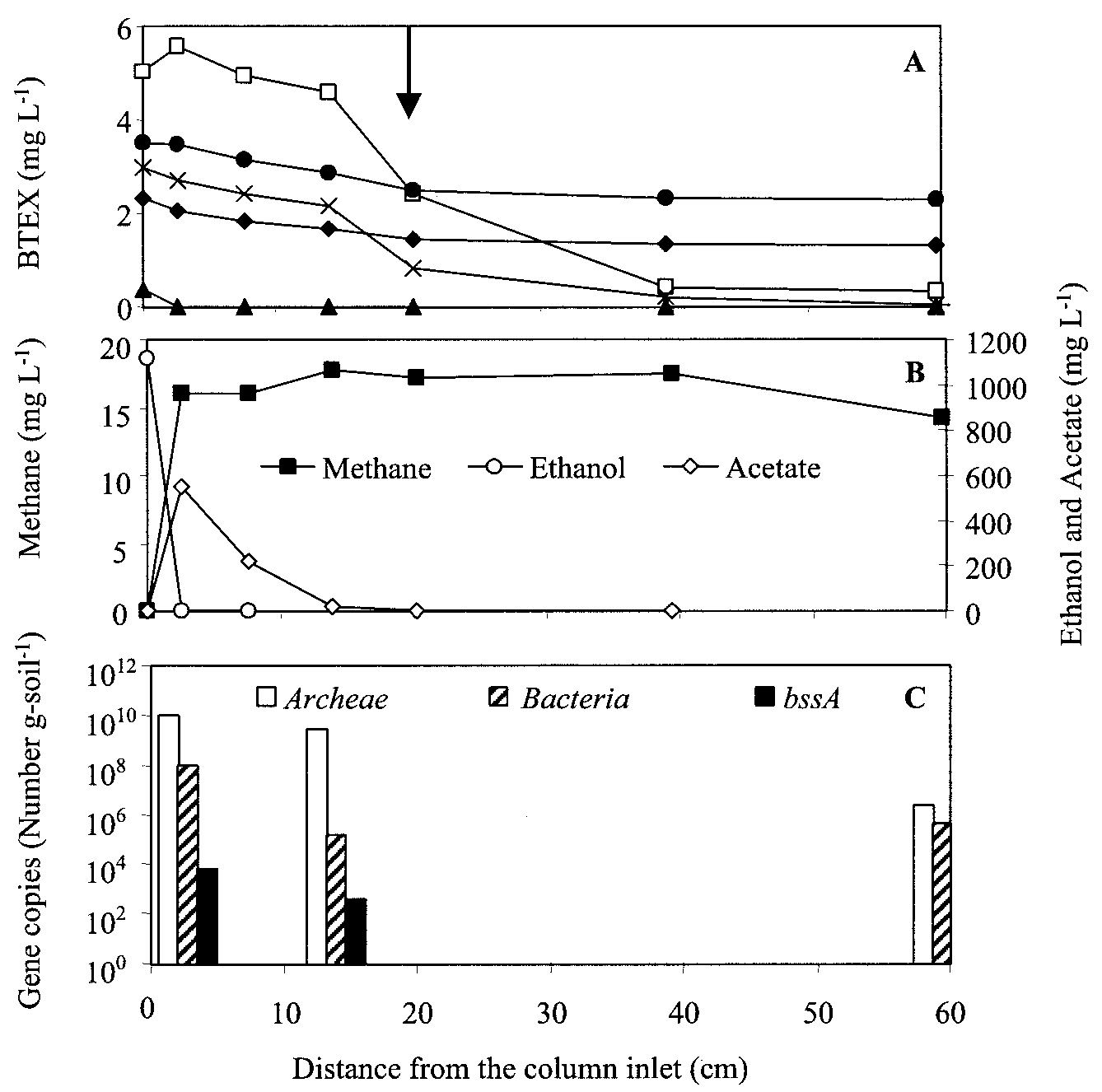
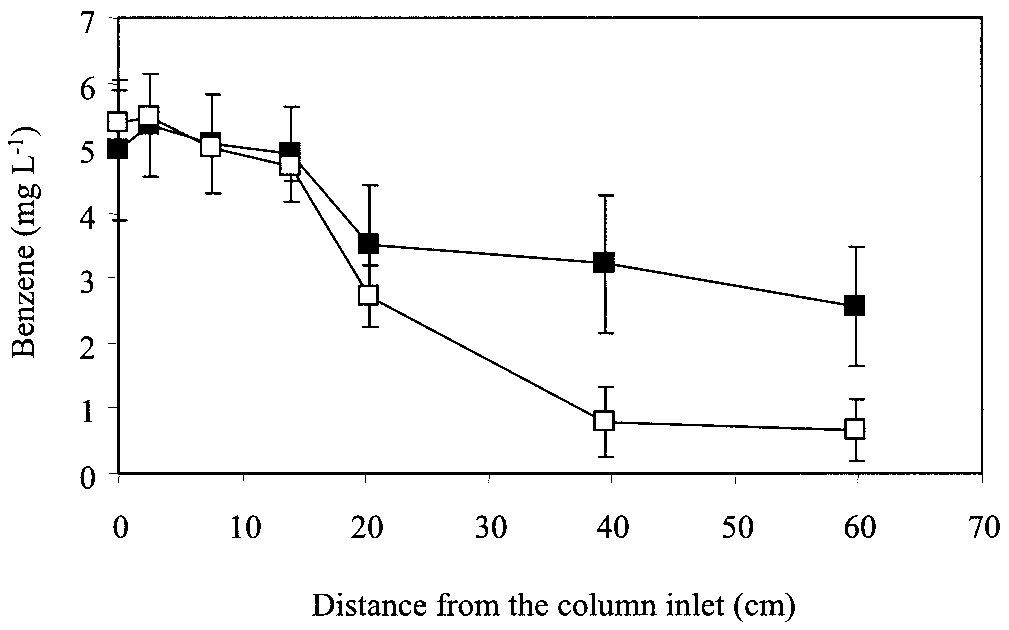 FIG. 3. Concentration profiles of the column bioaugmented with the benzene-enriched methanogenic consortium, taken 1 year after inocu-
lation. Concentrations of BTEX (A) and ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along the length
of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.
FIG. 3. Concentration profiles of the column bioaugmented with the benzene-enriched methanogenic consortium, taken 1 year after inocu-
lation. Concentrations of BTEX (A) and ethanol, acetate, and methane (B) and the bacterial gene distribution (C) are depicted along the length
of the column. The arrow shows the inoculation port. Symbols: ᮀ, benzene; Œ, toluene; }, ethylbenzene; ϫ, o-xylene; F, m-p-xylene.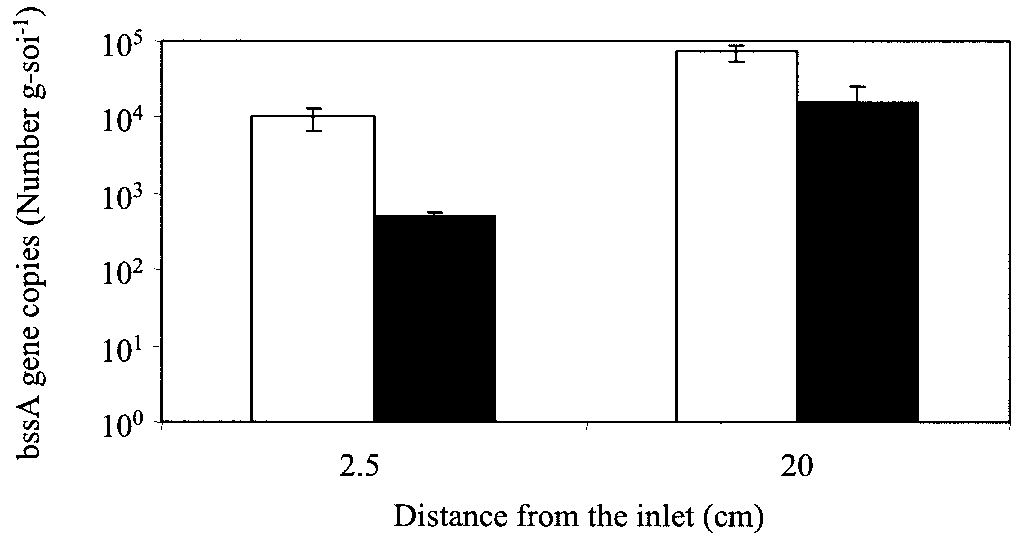 DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
bioaugmentation is unlikely to be universally applicable, and
pilot studies should be conducted to identify potential critical
limitations associated with scale-up issues, including the re-
quired inoculum size and cost, the need for pH control, and
performance at lower temperatures in the presence of poten-
ACKNOWLEDGMENTS
DEGRADATION OF BTEX-ETHANOL MIXTURES IN BIOAUGMENTED COLUMNS
bioaugmentation is unlikely to be universally applicable, and
pilot studies should be conducted to identify potential critical
limitations associated with scale-up issues, including the re-
quired inoculum size and cost, the need for pH control, and
performance at lower temperatures in the presence of poten-
ACKNOWLEDGMENTS