Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Somasimple.com
ATP-sensitive Kϩ channel blocker glibenclamideand diaphragm fatigue during normoxia and hypoxia
ERIK VAN LUNTEREN, MICHELLE MOYER, AND AUGUSTO TORRESDepartments of Medicine and Neurosciences, Case Western Reserve University,and Cleveland Veterans Affairs Medical Center, Cleveland, Ohio 44106Van Lunteren, Erik, Michelle Moyer, and Augusto
may potentially account for some of the discrepant
Torres. ATP-sensitive Kϩ channel blocker glibenclamide and
diaphragm fatigue during normoxia and hypoxia. J. Appl.
The purpose of the present study was to reexamine
Physiol. 85(2): 601–608, 1998.—The role of ATP-sensitive Kϩ
the issue of whether blocking KATP affects muscle
channels in skeletal muscle contractile performance is contro-
performance during repetitive contractions leading to
versial: blockers of these channels have been found to not
fatigue by 1) systematically addressing effects of stimu-
alter, accelerate, or attenuate fatigue. The present study
lation paradigm, temperature, and presence of hypoxia;
reexamined whether glibenclamide affects contractile perfor-
2) comparing intertrain with intratrain fatigue; and 3)
mance during repetitive contraction. Experiments systemati-
assessing the rate of muscle relaxation, which is known
cally assessed the effects of stimulation paradigm, tempera-
to slow during fatigue (10, 11, 16, 19, 26). We found that
ture, and presence of hypoxia and in addition comparedintertrain with intratrain fatigue. Adult rat diaphragm muscle
the KATP blocker glibenclamide significantly improves
strips were studied in vitro. At 37°C and normoxia, glibencla-
intratrain but not intertrain fatigue but only under
mide did not significantly affect any measure of fatigue
hypoxic and not normoxic conditions and that it slows
during continuous 5- or 100-Hz or intermittent 20-Hz stimu-
rate of muscle relaxation during fatigue under both
lation but progressively prolonged relaxation time during
normoxic and hypoxic conditions but not at low tempera-
20-Hz stimulation. At 20°C and normoxia, neither force nor
ture. These findings indicate that KATP may be acti-
relaxation rate was affected significantly by glibenclamide
vated during repetitive contraction, especially during
during 20-Hz stimulation. At 37°C and hypoxia, glibencla-
higher intensity contractions and/or under hypoxic
mide did not significantly affect fatigue at 5-Hz or intertrain
fatigue during 20-Hz stimulation but reduced intratrainfatigue and prolonged relaxation time during 20-Hz stimula-
tion. These findings indicate that, although ATP-sensitive Kϩchannels may be activated during repetitive contraction,
Male Sprague-Dawley rats (250–350 g) were anesthetized
their activation has only a modest effect on the rate of fatigue
with intraperitoneal urethan (1–1.5 g/kg), the diaphragmswere removed surgically, and two to four small strips (diam-
eter ϳ1–1.5 mm) were cut per animal, with care taken to
diaphragm; skeletal muscle; potassium; ATP-sensitive Kϩ
preserve the attachment of the muscle to the central tendon
and ribs. The muscle strips were mounted in physiologicalsolution at optimal length and were stimulated via platinumelectrodes by using a pulse width of 1 ms and supramaximalvoltages (Grass Instruments, West Warwick, RI). The aerated
2-5% CO2) physiological solution contained (in mM)
density in many tissues, including skeletal muscle (7).
135 NaCl, 5 KCl, 2.5 CaCl2, 1 MgSO4, 1 NaH2PO4, 15
In intact skeletal muscle fibers these channels are
NaHCO3, and 11 glucose, with the pH adjusted to 7.35–7.45. Bath temperature was controlled at 20 or 37°C by circulating
generally closed under resting normoxic conditions (2,
water of the appropriate temperature through the outer
3, 15). However, KATP open under conditions of low ATP
jacket of the tissue baths (Radnoti Glass, Monrovia, CA).
concentration ([ATP]), low intracellular pH, and meta-
Isometric tension was measured with a high-sensitivity trans-
bolic poisoning (2, 7, 9, 22). It has been postulated that
ducer (Kent Scientific/Radnoti Glass, Monrovia, CA). Twitch
KATP become activated during repetitive muscle contrac-
forces of ϳ0.5 kg/cm2 are obtained with this methodology in
tion especially during high-intensity contractions and/or
rat diaphragm (25). Force records were digitized, collected
under hypoxic stress, which thereby contributes to Kϩ
online with a computer (Axotape software, Axon Instruments,
efflux and the development of skeletal muscle fatigue
Foster City, CA), and stored for later data analysis. Drugs andreagents were obtained from Sigma Chemical (St. Louis,
MO). Glibenclamide was dissolved as a 2 mM stock solution
In support of the above postulate, studies utilizing
in 0.05 M NaOH, the proper volume of which was added to the
openers of KATP generally concur that these agents
bath to produce a final concentration of 100 µM (16).
accelerate fatigue, especially under hypoxic conditions
Diaphragm muscle strips were allowed to equilibrate and
(12, 28, 30). In contrast, studies utilizing blockers of
subsequently underwent twitch stimulation at 0.1 Hz for 3
min. Muscle strips were accepted for study only if twitch force
have found variable effects on fatigue: many
studies have found no significant effect on fatigue (5,
varied by no more than 5% during the 3-min baseline period.
12, 16, 28, 30), although an improvement (13, 14, 30)
Six separate experiments were performed, the conditions ofwhich are summarized in Table 1. Muscle strips were random-
and a worsening (6) of fatigue have also been reported.
ized across arms of a given experiment but not across
The methodology of these studies varies considerably
experiments. That is, muscle strips were randomized to
with respect to temperature, stimulation paradigm,
receive drug or no drug under a given set of experimental
presence of hypoxia, and data-analysis strategies, which
conditions; assignment of muscle strips was not randomized
8750-7587/98 $5.00 Copyright 1998 the American Physiological Society
Table 1. Stimulation frequencies, bath conditions,
All values presented are means Ϯ SE. Statistical analysis
of the effects of glibenclamide on prefatigue isometric twitch
and sample sizes of the six experiments
kinetics was performed with the unpaired t-test. Statistical
analysis of the effects of glibenclamide on peak force, force-330, and half relaxation times during fatigue runs was
performed with two-way ANOVA for repeated measures,
followed in the event of a significant ANOVA by the Newman-
Keuls test. The criterion for statistical significance was set at
Normoxic conditions (37°C). Under normoxic condi-
tions in nonfatigued muscle, glibenclamide (100 µM)did not significantly affect isometric twitch contractionor half relaxation times, although there was a nonsig-
across all six experimental conditions. Care was taken to
nificant trend for the latter to be prolonged (Table 2). In
ensure that muscle strips from a given animal were assigned
response to repetitive stimulation, glibenclamide did
to both drug and no drug. Each muscle strip was used onlyonce. Experiments A–C and E–F were conducted at 37°C,
not significantly affect peak force over time during 5-,
whereas experiment D was conducted at 20°C. After the
20-, or 100-Hz stimulation under normoxic conditions
baseline period, glibenclamide (100 µM) or vehicle (contain-
at 37°C (Fig. 1). Force-330, an evaluation of the ability
ing an equal volume of 0.05 M NaOH) was added to the bath
of the muscle to maintain force during the plateau
for all experiments, which was followed by an equilibration
phase within the same tetanic stimulation (evaluated
period of 4 min. For experiments E–F only, the gas with which
during 20-Hz trains), was slightly but not significantly
the solution was aerated was subsequently switched to 95%
improved by glibenclamide under normoxic conditions
N2-5% CO2 followed by an equilibration period of 4 min. Bath
(Fig. 2). However, the extent to which the half relax-
oxygen tension was measured in some of the hypoxia studies
ation time progressively prolonged during repetitive
with a dissolved-oxygen meter (ISO-2, World Precision Instru-
stimulation was augmented significantly by glibencla-
ments, Sarasota, FL) and averaged 3.8 Ϯ 0.8% at the end ofthe 4-min equilibration period. The muscles were stimulated
mide during 20-Hz and to a lesser extent 5-Hz stimula-
at 0.1 Hz to monitor twitch tension during all of the above.
Finally, the muscle strips underwent one of three stimulation
Effects of lowering temperature (normoxia). Effects of
paradigms: continuous 5-Hz stimulation (experiments A and
lowering temperature to 20°C were assessed during
E), intermittent 20-Hz stimulation (train duration 0.33 s,
fatigue produced by 20-Hz stimulation. At this tempera-
with 1 train delivered every second) (experiments B, D, and
ture, glibenclamide had no significant effect on baseline
F), or continuous 100-Hz stimulation (experiment C). Only
twitch kinetics (Table 2), peak force over time (Fig. 4,
limited studies were done at 20°C because it would be highly
left), force-330 (Fig. 4, middle), or rate of relaxation
unusual for mammalian muscle to be contracting in vivo at
this cold a temperature, whereas tissue hypoxia and alter-
Effects of hypoxia (37°C). Glibenclamide had no sig-
ations in motoneuronal firing frequency can be seen under avariety of circumstances.
nificant effect on isometric twitch contraction and
Force records were analyzed offline with use of manually
half relaxation times under hypoxic conditions (Table
positioned cursors displayed on the computer screen. Isomet-ric tension was measured in grams and subsequently normal-ized for each muscle strip to the average of the last three
Table 2. Effects of glibenclamide on diaphragm
twitches during the (predrug) baseline period. Normalization
isometric twitch contraction and half relaxation
was performed to minimize the confounding effects of inter-
strip variability in size and hence baseline force and to reducethe influences of slight variations in dissection technique
affecting baseline force. This method of normalization isconsistent with approaches used by us (24, 25, 27) and others
(30) in studies of Kϩ channel blockers. Other studies of KATP
blockers have normalized force to postdrug, prefatigue values
(e.g., 13, 16, 28), which is similar to the present approach in
ATP blockers in the concentrations used generally have
minimal effects on baseline force. Intratrain fatigue was
assessed during 20-Hz stimulation by measuring the force at
the end of the 330-ms-long train and expressing this as a
percentage of the maximum force within the same tetanus
(force-330) (26; as modified from Ref. 16). During 0.1- and
5-Hz stimulation, contraction time was assessed as the
amount of time for twitch force to reach its peak, and
Values are means Ϯ SE from 0.1-Hz twitches immediately preced-
half relaxation time was assessed as the amount of time for
ing onset of the fatiguing stimulation. Values are from experiments
twitch force to decay to one-half of the peak value. During
A–C for normoxia at 37°C, experiment D for normoxia at 20°C, and
20-Hz trains, contraction time was assessed from the first
experiments E and F for hypoxia at 37°C. Force was normalized to the
twitch of the train, and relaxation time was assessed from the
value for twitch force immediately before addition of drug or no drug
decay in force at the end of the train.
Fig. 1. Changes in peak diaphragm force over timeduring repetitive 5- (A), 20- (B), and 100-Hz (C) stimula-tion in presence and absence of glibenclamide (100 µM)under normoxic conditions and a temperature of 37°C. Force values are means Ϯ SE and are normalized to thevalue for twitch force immediately before addition ofdrug or no drug as described in METHODS. There were nosignificant effects of glibenclamide at 5 Hz (P ϭ 0.41), 20Hz (P ϭ 0.24) or 100 Hz (P ϭ 0.75).
2). The change in peak force over time during 5- and
DISCUSSION
20-Hz stimulation was not affected significantly by
Methodological issues. There are a number of agents
glibenclamide under hypoxic conditions, although there
was a trend for peak force to be improved by glibencla-
mide during 20-Hz stimulation (Fig. 5). In contrast to
previous studies of muscle contractility. Gibenclamide
during normoxia, force-330 (assessed during 20-Hz
was chosen for the present study on the basis of two
trains) was improved significantly by glibenclamide
major considerations. First, glibenclamide at the concen-
during hypoxia (Fig. 6). The extent to which relaxation
tration used in this study (100 µM) blocks rat skeletal
rate slowed during repetitive 20-Hz stimulation was
muscle KATP but not voltage-gated Kϩ channels or
generally more prominent during hypoxia than during
Ca2ϩ-activated Kϩ channels (17); comparable data on
normoxia, and this was augmented significantly by
sensitivity and specificity in rat skeletal muscle are not
glibenclamide during 20- but not 5-Hz stimulation
available for the other KATP blockers. However, tolbuta-
mide affects muscle excitability, suggesting that it mayhave effects in addition to blocking KATP (5). Thus it wasfelt best to pick the agent for which specificity for KATPwas best established in rat skeletal muscle. Second,glibenclamide has been used in the majority of previousstudies examining muscle fatigue and KATP. Amongeight studies, six used glibenclamide (6, 12–14, 16, 30),two used glyburide (5, 28), and one study each usedphentolamine (30), ciclazindol (30), and tolbutamide(5). It is easier to compare the present data with otherdata by choosing the agent used most commonly inprevious studies (glibenclamide) rather than anotheragent (e.g., tolbutamide, glyburide, ciclazinol).
Light and French (17) examined the sensitivity to
glibenclamide of KATP reconstituted from rat skeletalmuscle. They noted a concentration for one-half inhibi-tion of open probability (Ki) of 3–5 µM and found that adose of 10–100 µM was sufficient to fully eliminatevisible channel openings. The value for Ki in rat muscleis higher than that of mouse muscle (Ki of 190 nM) (1). Light and French (17) also found that 100 µM glibencla-
Fig. 2. Effects of glibenclamide (100 µM) on ability of diaphragm to
mide had no effects on voltage-gated or Ca2ϩ-activated
maintain force during plateau phase within the same tetanic stimula-
Kϩ channels, suggesting good specificity for K
tion (force-330) during 20-Hz stimulation under normoxic conditions
and a temperature of 37°C. Values are means Ϯ SE. Force-330, a
sistent with studies in other tissues). A glibenclamide
measure of intratrain fatigue, was determined by evaluating force of
concentration of 100 µM was chosen over 10 µM in the
the last contraction in the train as a percentage of the maximum
present study for several reasons. First, a concentra-
tetanic force of the same tetanic contraction at each time point. There
tion of glibenclamide was desired that would definitely
was a nonsignificant trend for glibenclamide to improve force-330(P ϭ 0.12).
block KATP so that any absent effects of glibenclamide
Fig. 3. Diaphragm half relaxation time over the courseof repetitive 5- (A) and 20-Hz (B) stimulation in thepresence and absence of glibenclamide (100 µM) undernormoxic conditions and a temperature of 37°C. Valuesare means Ϯ SE. *Significant differences between gliben-clamide and no drug, P Ͻ 0.05.
on fatigue could not be attributed to a concentration
clamide and glyburide in either DMSO (28, 30) or
that was possibly too low. Second, the present study
NaOH (6, 16). The latter was chosen for the present
used muscle strips, whereas French and Light (17)
study because DMSO affects free radicals and thereby
studied biplanar layers; a concentration higher than
muscle contractile performance and, hence, could have
the minimal amount needed would ensure that an
greater confounding effects than a slight increase in pH
adequate concentration of drug would reach the center
induced by NaOH. Control and drug-treated muscle
of the muscle strip. Third, a concentration of 100 µM
strips had an equal amount of NaOH added to the bath
was used in the most detailed previous study of gliben-
so that any potential effects of acid-base changes would
clamide and muscle fatigue (which also included data
on action potentials) (16) so that direct comparisons
An incubation period for glibenclamide of 4 min was
could most easily be made by using the same drug
used in the present study. Light et al. (16) examined
effects of glibenclamide (100 µM) on muscle action
In the present study, glibenclamide was dissolved in
potential repolarization at a temperature of 20°C. They
NaOH as a stock solution before it was added to the
found that action potential repolarization was slowed
bath. Glibenclamide (and glyburide) do not dissolve
by fatigue and that glibenclamide further slowed action
readily in water or saline. Previous studies of KATP
potential repolarization. Furthermore, the mean val-
blockers and muscle contraction have dissolved gliben-
ues of the half repolarization time after fatigue in the
Fig. 4. Effect of glibenclamide (100 µM) on diaphragmduring 20-Hz stimulation under normoxic conditionsand a temperature of 20°C. Values are means Ϯ SE. Changes in peak force (left), force-330 (middle), and halfrelaxation time (right) are indicated. Peak force wasnormalized to the value for force immediately beforeaddition of drug or no drug, as described in METHODS. Glibenclamide had no significant effects on peak force(P ϭ 0.81), force-330 (P ϭ 0.94), or half relaxation time(P ϭ 0.36).
Fig. 5. Alterations in peak diaphragm force over timeduring repetitive 5- (A) and 20-Hz (B) stimulation inpresence and absence of glibenclamide (100 µM) underhypoxic conditions and a temperature of 37°C. Forcevalues are means Ϯ SE and are normalized to the valuefor twitch force immediately before addition of drug orno drug as described in METHODS. There were no signifi-cant effects of glibenclamide during 5-Hz stimulation,but there was a nonsignificant trend for force to improveduring 20-Hz stimulation (P ϭ 0.07).
presence of glibenclamide were the same whether the
assessed by utilizing a variety of stimulation frequen-
drug was applied 60 min before fatigue or 60 s before
cies, ranging from 0.2 to 140 Hz (5, 6, 12, 13, 16, 28, 30).
the end of fatigue. The latter data suggest a fast rate of
In addition, one study used a spontaneously breathing
diffusion and a fast onset of action of glibenclamide
model to test diaphragm fatigue (14), in which motoneu-
(Յ60 s) in skeletal muscle tissue. Light et al. studied
ronal firing frequency was not assessed but would be
muscle fiber bundles with diameters of 1–1.5 mm,
expected to vary among motor units and over time.
which is the same size used in the present study. They
During muscle contraction, Kϩ efflux and Naϩ influx
used a temperature of 20°C, whereas the present study
and the resultant alteration in transmembranous Kϩ
used a temperature of 37°C; diffusion and onset of drug
and Naϩ concentration gradients may lead to sarcolem-
action should be faster at the higher temperature.
mal depolarization, especially in the T tubules in which
Based on these data, 4 min should be sufficiently long
diffusion of ions is slower than at the outer surface of
for equilibration after drug addition.
the muscle (21, 29). Much of the Kϩ efflux during
Effects of glibenclamide on muscle contraction. Ef-
fects of KATP blockers on muscle fatigue have been
Fig. 7. Diaphragm half relaxation time over the course of repetitive
Fig. 6. Effects of glibenclamide (100 µM) on ability of diaphragm to
5- (A) and 20-Hz (B) stimulation in presence and absence of glibencla-
maintain force during plateau phase within same tetanic stimulation
mide (100 µM) under hypoxic conditions and a temperature of 37°C.
(force-330) during 20-Hz stimulation under hypoxic conditions and a
Values are means Ϯ SE. Half relaxation times were difficult to
temperature of 37°C. Values are means Ϯ SE. Force-330, a measure
quantify accurately when force values became very small toward the
of intratrain fatigue, was determined by evaluating force of the last
end of fatiguing stimulation (see Fig. 5) and hence are reported only
contraction in the train as percentage of maximum tetanic force of the
for the first 2 min of the 3-min stimulation period. * Significant
same tetanic contraction at each time point. * Significant differences
differences between glibenclamide and no drug, P Ͻ 0.05. Contrac-
between glibenclamide and no drug, P Ͻ 0.05.
tion time was not affected significantly by glibenclamide (P ϭ 0.39).
contraction occurs via delayed rectifier Kϩ channels
Light et al., we found a slowing of relaxation rate by
(29), although KATP has been postulated to contribute to
glibenclamide during fatigue produced during 20-Hz
the Kϩ efflux under conditions of depleted intracellular
stimulation under both normoxic and hypoxic condi-
[ATP] and concommitant acidosis (8, 21). The Naϩ-Kϩ-
tions. These data suggest that KATP may be activated
ATPase will restore the membranous ion gradients
during fatiguing stimuli but to an insufficient extent to
back to normal, but at high rates of muscle contraction
affect peak force production. This is consistent with a
the active transport is overwhelmed (4). Thus the role
previously proposed explanation for glibenclamide not
of Kϩ channels in regulating muscle fatigue is believed
affecting fatigue but delaying the recovery from fatigue
to be most prominent during intense muscle activation.
Hence Kϩ channel blockers should improve high-
The mechanism by which altering Kϩ channel conduc-
frequency more than low-frequency fatigue and intra-
tance affects muscle relaxation rate is unlikely to be a
train more than intertrain fatigue. That one of the
direct effect on either the rate of Ca2ϩ reuptake by the
studies with the greatest beneficial effects of KATP
sarcoplasmic reticulum or the rate of Ca2ϩ binding by
blockade on fatigue used a low stimulation rate of 0.25
parvalbumin. More likely, the effects of Kϩ channels on
Hz (13) is therefore surprising and may very well reflect
the rate of relaxation are mediated by altering the rate
other methodological differences (e.g., use of an in vivo
of action potential repolarization. Normally, membrane
preparation in which vascular or other systemic effects
potential repolarizes very quickly during an action
of glibenclamide may have contributed to the findings)
potential. As a result, there is only a brief period of time
compared with the other studies of KATP blockers (5, 6,
during repolarization when there is continued Ca2ϩ
influx while Ca2ϩ is simultaneously being taken back
In the present study we found no significant effects of
up by the sarcoplasmic reticulum and/or being bound to
glibenclamide on fatigue during continuous 5- or 100-Hz
intracellular Ca2ϩ buffers. If action potential repolariza-
stimulation or on intertrain fatigue during intermit-
tion is slowed (e.g., with Kϩ channel blockade), this
tent 20-Hz stimulation under normoxic or hypoxic
period can be prolonged, thereby slowing the rate at
conditions, consistent with all of the other in vitro
which intracellular Ca2ϩ concentration ([Ca2ϩ]) falls
studies of glibenclamide and fatigue (5, 12, 16, 28, 30).
and hence slowing the rate of relaxation. If the degree
The only in vitro study reporting an improvement of
of action potential repolarization slowing is small, it
intertrain fatigue with KATP blockers noted a modest
may not be sufficient to affect mechanical relaxation.
improvement in fatigue with ciclazinol but not with
This could explain why Light et al. (16) found action
glibenclamide (30), suggesting that ciclazinol may be a
potential prolongation but no slowing of relaxation
more effective blocker of KATP or may have additional
with glibenclamide. On the other hand, if the degree of
effects in addition to blocking KATP (e.g., blocking other
action potential slowing is large, either contraction or
Kϩ channels). In the present study we found no signifi-
relaxation time could be slowed depending on the
cant effects of glibenclamide on intratrain fatigue dur-
kinetics of the changes in intracellular [Ca2ϩ] relative
ing normoxia (although there was a trend toward
to the kinetics of actin-myosin interactions. As muscle
improvement), but we found an attenuation of intra-
fatigues, action potential repolarization slows and relax-
train fatigue during hypoxia. The former finding (nor-
ation rate slows. Under these circumstances, effects of
moxic conditions) is consistent with two previous stud-
Kϩ channel blockers on rate of relaxation may become
ies (5, 16), neither of which, however, examined
more manifest, as was found for glibenclamide in the
intratrain fatigue under hypoxic conditions. The pres-
present study. This is consistent with previous studies
ent finding of glibenclamide significantly attenuating
of the Kϩ channel-blocking aminopyridines, which do
only intratrain fatigue and only during hypoxia sug-
not slow relaxation rate in nonfatigued muscle but
gests that the contribution of KATP to fatigue is small
markedly augment slowing of relaxation rate as muscle
and is limited to conditions expected to lead to profound
undergoes fatiguing contractions (25, 26).
ATP depletion and/or intracellular acidosis.
Effects of KATP blockers have been assessed at 20°C
The rate of muscle relaxation slows with fatigue and
(5, 16), 30°C (28), or 37–38°C (6, 12–14, 30). Studies at
especially does so under hypoxic conditions (10, 11, 16,
20°C utilized frog muscle, whereas studies performed
19, 26). Two previous studies have found that glyburide
at 30–38°C utilized mammalian muscle so that the
and glibenclamide slow the rate of action potential
influence of temperature on muscle contractile re-
repolarization in resting and fatigued muscle (5, 16).
sponses to KATP blockers cannot be inferred directly
Surprisingly, the rate of muscle relaxation was not
from previous work. Of note, however, is that both of
found to be affected by glibenclamide in either resting
the studies at a cool temperature found no effect of KATP
or fatigued muscle in a previous study despite changes
blockers on fatigue, whereas the studies at warmer
in action potential repolarization rate (16). Data on
temperatures have noted variable effects of KATP block-
muscle relaxation rate were not provided for glyburide
ers on fatigue. Ion channels are very sensitive to
(5), nor have other studies of glibenclamide and fatigue
temperature, with rates of activation and deactivation
reported values for rate of muscle relaxation. The
having especially high values for Q10 compared with
present data concur with those of Light et al. (16), who
peak current; furthermore, values for Q10 may vary as a
found that that KATP blockade does not significantly
function of membrane potential (see, e.g., Refs. 20, 23).
alter rate of relaxation of resting muscle. In contrast to
In the present study of 20-Hz stimulation during
normoxia, we found that at neither warm nor cold
Address for reprint requests: E. van Lunteren, Pulmonary Sect.
temperature was there a significant effect of glibencla-
111J(W), Cleveland VA Medical Center, 10701 East Boulevard,Cleveland, OH 44106 (E-mail: exv4@po.cwru.edu).
mide on either intertrain or intratrain fatigue. Thissuggests that differences among previous studies re-
Received 12 June 1997; accepted in final form 1 April 1998.
garding whether fatigue is attenuated with glibencla-
REFERENCES
mide are unlikely to be due to differences in muscle
1. Allard, B., and M. Lazdunski. Pharmacological properties of
temperature. On the other hand, we found that, during
ATP-sensitive Kϩ channels in mammalian skeletal muscle. Eur. J.
fatiguing 20-Hz stimulation, glibenclamide led to an
Pharmacol. 236: 419–426, 1993.
augmentation of the relaxation rate prolongation at
2. Allard, B., M. Lazdunski, and O. Rougier. Activation of
37°C but not at 20°C. Therefore, a low temperature
ATP-dependent Kϩ channels by metabolic poisoning in adultmouse skeletal muscle: role of intracellular Mg2ϩ and pH. J.
may explain why Light et al. (16), having conducted
Physiol. (Lond.) 485: 283–296, 1995.
their studies at 20°C, did not find that glibenclamide
3. Brinkmeier, H., E. Zachar, and R. Rudel. Voltage-dependent
prolonged relaxation rate during fatigue despite a high
Kϩ channels in the sarcolemma of mouse skeletal muscle. ¨ gers Arch. 419: 486–491, 1991.
4. Clausen, T. Regulation of active Naϩ-Kϩ transport in skeletal
Pharmacological agents that block many types of Kϩ
muscle. Physiol. Rev. 66: 542–580, 1986.
channels (e.g., tetraethylammonium, 4-aminopyridine,
5. Comtois, A., P. Light, J. M. Renaud, and M. K. Kong.
3,4-diaminopyridine) have greater effects on muscle
Tolbutamide, but not glyburide, affects the excitability and
contractile performance than have been noted for the
contractility of unfatigued frog sartorius muscle. Eur. J. Pharma-
6. Comtois, A., C. Sinderby, N. Comtois, A. Grassino, and J. M.
force of resting muscle range from ϳ10 to 150% with
Renaud. An ATP-sensitive potassium channel blocker decreased
single agents and several hundred percent with combi-
diaphragmatic circulation in anesthetized dogs. J. Appl. Physiol.
nations of agents, and these increases are accompanied
by prolongations of contraction time and for the amino-
7. Davies, N. W. Modulation of ATP-sensitive Kϩ channels in
skeletal muscle by intracellular protons. Nature 343: 375–377,
pyridines a leftward shift in the force-frequency relation-
ship (18, 24, 25, 27). Furthermore, the aminopyridines
8. Davies, N. W., N. B. Standen, and P. R. Stanfield. ATP-
improve both peak force and maintenance of intratrain
dependent potassium channels of muscle cells: their properties,
force during repetitive 20-Hz stimulation, reduce the
regulation, and possible functions. J. Bioenerg. Biomembr. 23:509–535, 1991.
neurotransmission failure contribution to fatigue, and
9. Davies, N. W., N. B. Standen, and P. R. Stanfield. The effect of
augment the slowing of relaxation rate during fatigue
intracellular pH on ATP-dependent potassium channels of frog
(all tested under normoxic conditions) (24, 25). There-
skeletal muscle. J. Physiol. (Lond.) 445: 549–568, 1992.
10. Dawson, M. J., D. G. Gadian, and D. R. Wilkie. Mechanical
relaxation rate and metabolism studied in fatiguing muscle by
compared with other Kϩ channels (e.g., delayed recti-
phosphorus nuclear magnetic resonance. J. Physiol. (Lond.) 299:
fier Kϩ channels) in regulating muscle contractile per-
formance. However, there may be other conditions
11. Esau, S. A., F. Bellemare, A. Grassino, S. Permutt, C. Rous- sos, and R. L. Pardy. Changes in relaxation rate with diaphragm
fatigue in humans. J. Appl. Physiol. 54: 1353–1360, 1983.
modulating muscle force. Specifically, ATP consump-
12. Gurden, J. M., and V. J. Hart. Effects of cromakalim and
tion may be higher during repetitive dynamic contrac-
glibenclamide on rabbit isolated tenuissimus muscle during
tions (contractions associated with muscle shortening)
hypoxia or normoxia (Abstract). Br. J. Pharmacol. 114: 237P,
than during isometric contractions, which could lead to
13. Gutierrez, G., G. Arbelaez, R. A. Cabezas, and D. Lee.
a greater fall in intracellular [ATP] and hence a greater
Glibenclamide ameliorates skeletal muscle fatigue (Abstract).
activation of KATP. To test this, future studies are
Am. J. Respir. Crit. Care Med. 151: A811, 1995.
needed to determine the extent to which glibenclamide
14. Gutierrez, G., D. Lee, and R. Cabezas. Glibenclamide pre-
modulates muscle shortening velocity during fatiguing
vents rabbit diaphragmatic fatigue (Abstract). Am. J. Respir. Crit. Care Med. 153: A687, 1996.
15. Hussain, M., A. C. Wareham, and S. I. Head. Mechanism of
In conclusion, KATP contributes to fatigue production
action of a Kϩ channel activator BRL 38227 on ATP-sensitive Kϩ
during isometric contractions only under extreme con-
channels in mouse skeletal muscle fibers. J. Physiol. (Lond.) 478:
ditions, such as intense stimulation during hypoxia.
16. Light, P. E., A. S. Comtois, and J. M. Renaud. The effect of
These channels are activated under less extreme condi-
glibenclamide on frog skeletal muscle: evidence for KϩATP chan-
tions as evidenced by a prolonged relaxation time as
nel activation during fatigue. J. Physiol. (Lond.) 475: 495–507,
fatigue develops, but the degree of activation is not
sufficient to substantially decrease force production.
17. Light, P. E., and R. J. French. Glibenclamide selectively blocks
Intense stimulation during hypoxia may result in an
ATP-sensitive Kϩ channels reconstituted from skeletal muscle. Eur. J. Pharmacol. 259: 219–222, 1994.
acidic intracellular environment that is partially de-
18. Lin-Shiau, S. Y., S. Y. Day, and W. M. Fu. Use of ion channel
pleted of ATP. Under these circumstances, ATP-
blockers in studying the regulation of skeletal muscle contrac-
dependent Naϩ-Kϩ pumps are no longer sufficient to
tions. Naunyn Schmiedebergs Arch. Pharmacol. 344: 691–697,
restore normal transmembrane ion gradients so that
19. Metzger, J. M., and R. H. Fitts. Fatigue from high- and
Kϩ efflux through KATP may contribute to fatigue.
low-frequency muscle stimulation: role of sarcolemma actionpotentials. Exp. Neurol. 93: 320–333, 1986.
This study was supported in part by the Veterans Affairs Medical
20. Nobile, M., E. Carbone, H. D. Lux, and H. Zucker. Tempera-
Research Service and by National Heart, Lung, and Blood Institute
ture sensitivity of Ca currents in chick sensory neurons. Pflu
Specialized Center of Research Grant HL-42215.
21. Sjogaard, G. Role of exercise-induced potassium fluxes underly-
26. Van Lunteren, E., A. Torres, and M. Moyer. Effects of hypoxia
ing muscle fatigue: a brief review. Can. J. Physiol. Pharmacol.
on diaphragm relaxation rate during fatigue. J. Appl. Physiol.
22. Spruce, A. E., N. B. Standen, and P. R. Stanfield. Voltage-
27. Van Lunteren, E., H. Vafaie, and M. Moyer. Changes in
dependent ATP-sensitive potassium channels of skeletal muscle
pharyngeal respiratory muscle force produced by Kϩ channel
membrane. Nature 316: 736–738, 1985.
blockade. Respir. Physiol. 99: 331–340, 1995.
23. Van Lunteren, E., K. S. Elmslie, and S. W. Jones. Effects of
28. Wesselcouch, E. O., C. Sargent, M. W. Wilde, and M. A.
temperature on calcium current of bullfrog sympathetic neurons. Smith. ATP-sensitive potassium channels and skeletal muscle J. Physiol. (Lond.) 466: 81–93, 1993.
function in vitro. J. Exp. Pharmacol. Ther. 267: 410–416, 1993.
24. Van Lunteren, E., and M. Moyer. Effects of 3,4-diaminopyri-
29. Westerblad, H., J. A. Lee, J. Lannergren, and D. G. Allen.
dine on diaphragm force and fatigue, including fatigue due to
Cellular mechanisms of fatigue in skeletal muscle. Am. J.
neurotransmission failure. J. Appl. Physiol. 81: 2214–2220,
Physiol. 261 (Cell Physiol. 30): C195–C209, 1991.
30. Wickendon, A. D., H. Prior, E. Kelly, K. Russell, S. M.
25. Van Lunteren, E., M. Moyer, and A. Torres. Effect of Kϩ Poucher, and P. Kumar. The effects of pharmacological modu-
channel blockade on fatigue in rat diaphragm muscle. J. Appl.
lation of KATP on the guinea-pig isolated diaphragm. Eur. J.Physiol. 79: 738–747, 1995. Pharmacol. 302: 79–88, 1996.
Bangladesch-Rundbrief Nr. 48 Chronik 1 BANGLADESCH-CHRONIK Menschenrechte: Die Sicherheitskräfte verhindern einen Sturm der islamistischen Anhänger der International Khatme Nabuwat Bewegung auf eine Moschee der Ahmadiyya-Gemeinde in Dhanikhola im Distrikt Mymensingh. Protestaktionen gegen Ahmadiyya-Moscheen in der Provinz Sylhet (24. März) und in Shyhamnager in der Provinz Satkhira (12.
 ATP-sensitive Kϩ channel blocker glibenclamideand diaphragm fatigue during normoxia and hypoxia
ERIK VAN LUNTEREN, MICHELLE MOYER, AND AUGUSTO TORRESDepartments of Medicine and Neurosciences, Case Western Reserve University,and Cleveland Veterans Affairs Medical Center, Cleveland, Ohio 44106
Van Lunteren, Erik, Michelle Moyer, and Augusto
ATP-sensitive Kϩ channel blocker glibenclamideand diaphragm fatigue during normoxia and hypoxia
ERIK VAN LUNTEREN, MICHELLE MOYER, AND AUGUSTO TORRESDepartments of Medicine and Neurosciences, Case Western Reserve University,and Cleveland Veterans Affairs Medical Center, Cleveland, Ohio 44106
Van Lunteren, Erik, Michelle Moyer, and Augusto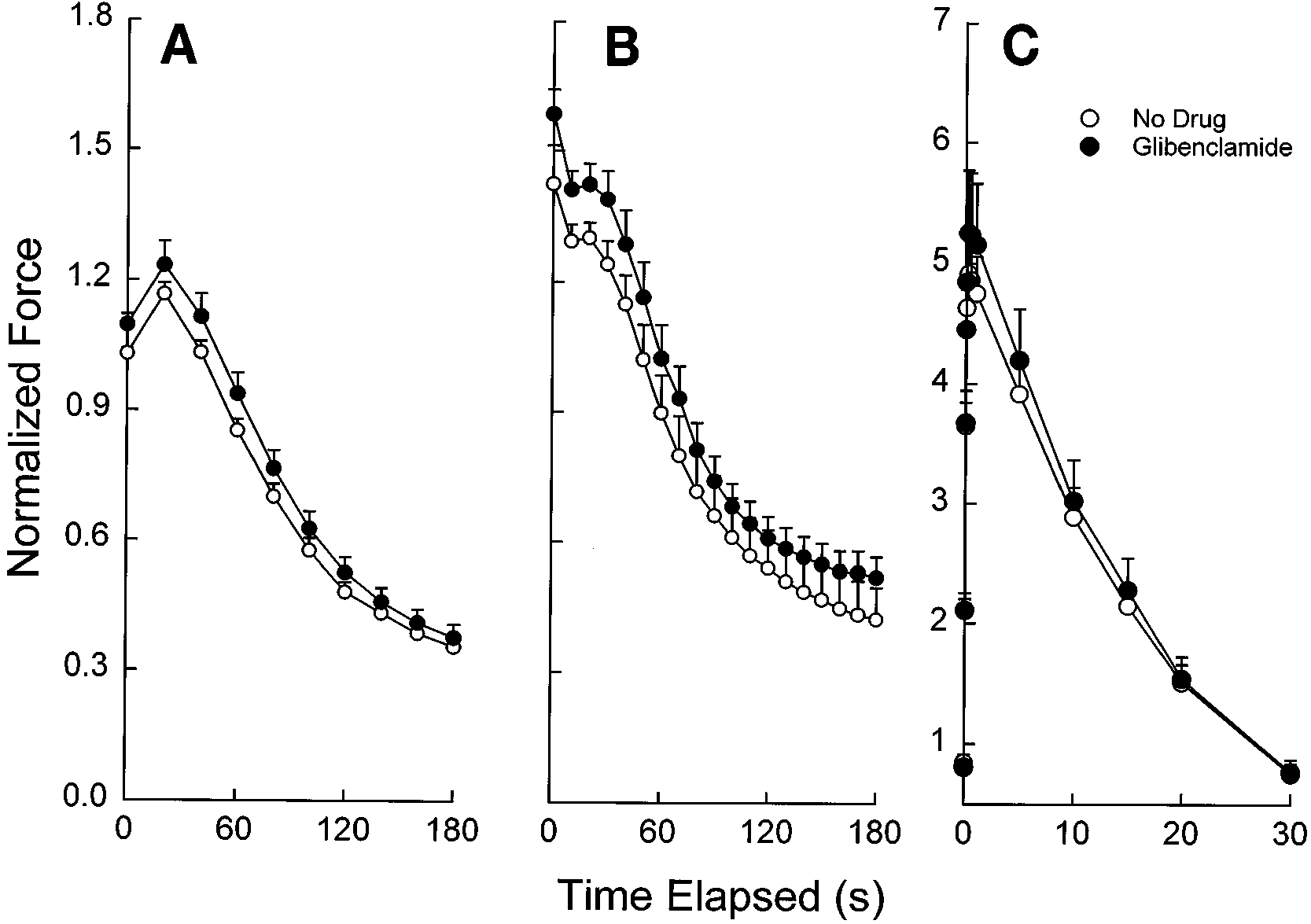
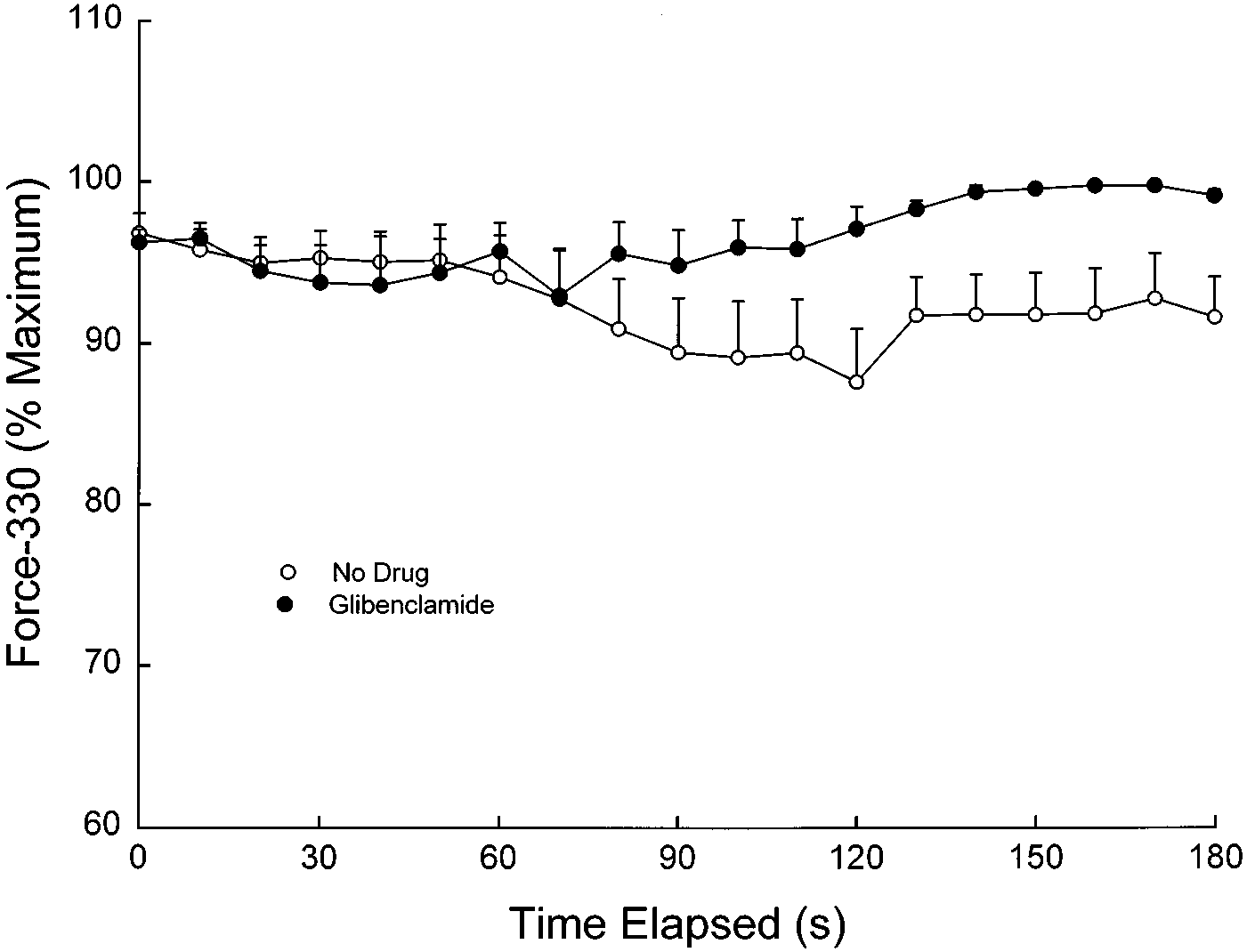 Fig. 1. Changes in peak diaphragm force over timeduring repetitive 5- (A), 20- (B), and 100-Hz (C) stimula-tion in presence and absence of glibenclamide (100 µM)under normoxic conditions and a temperature of 37°C.
Fig. 1. Changes in peak diaphragm force over timeduring repetitive 5- (A), 20- (B), and 100-Hz (C) stimula-tion in presence and absence of glibenclamide (100 µM)under normoxic conditions and a temperature of 37°C.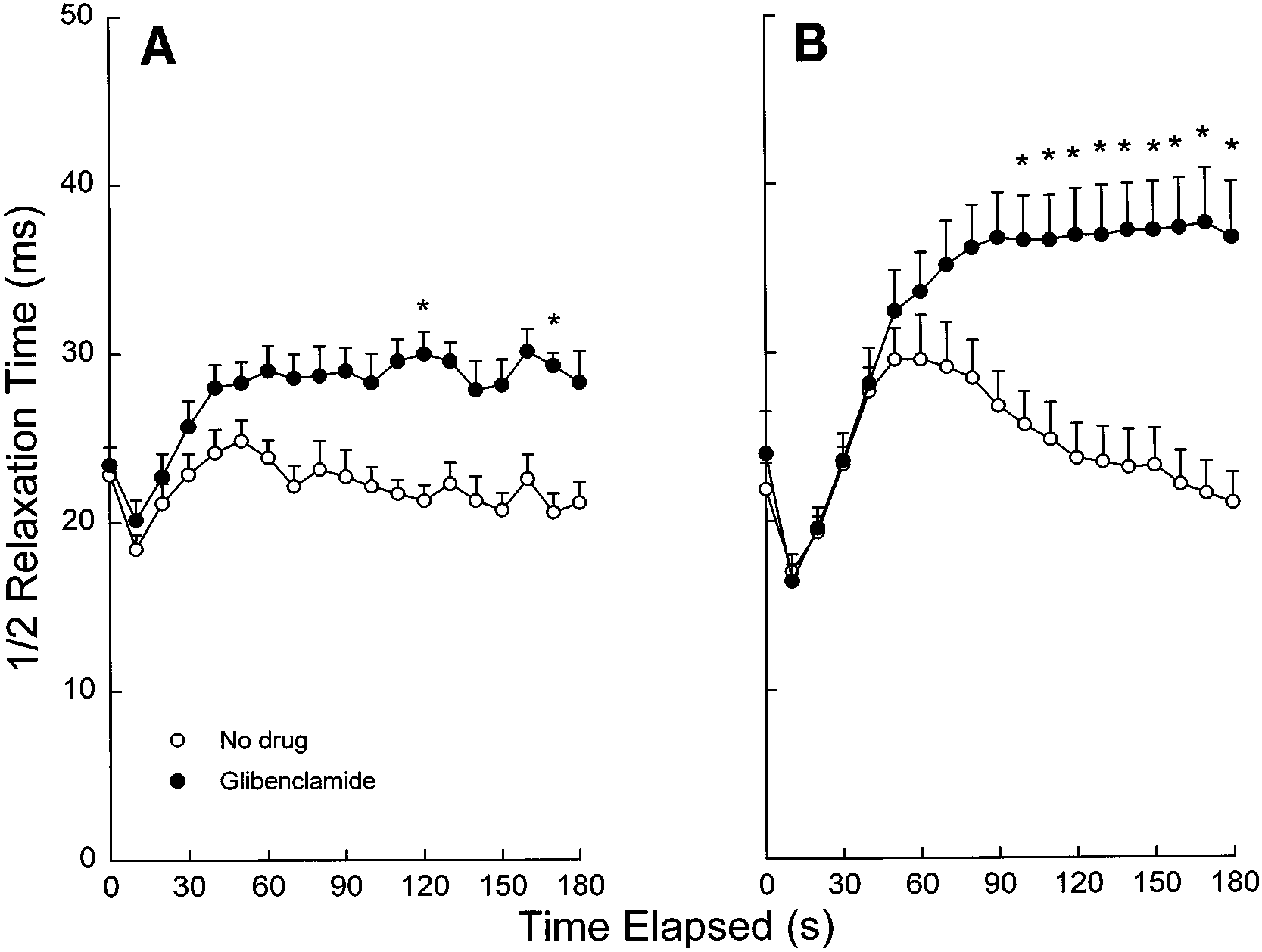
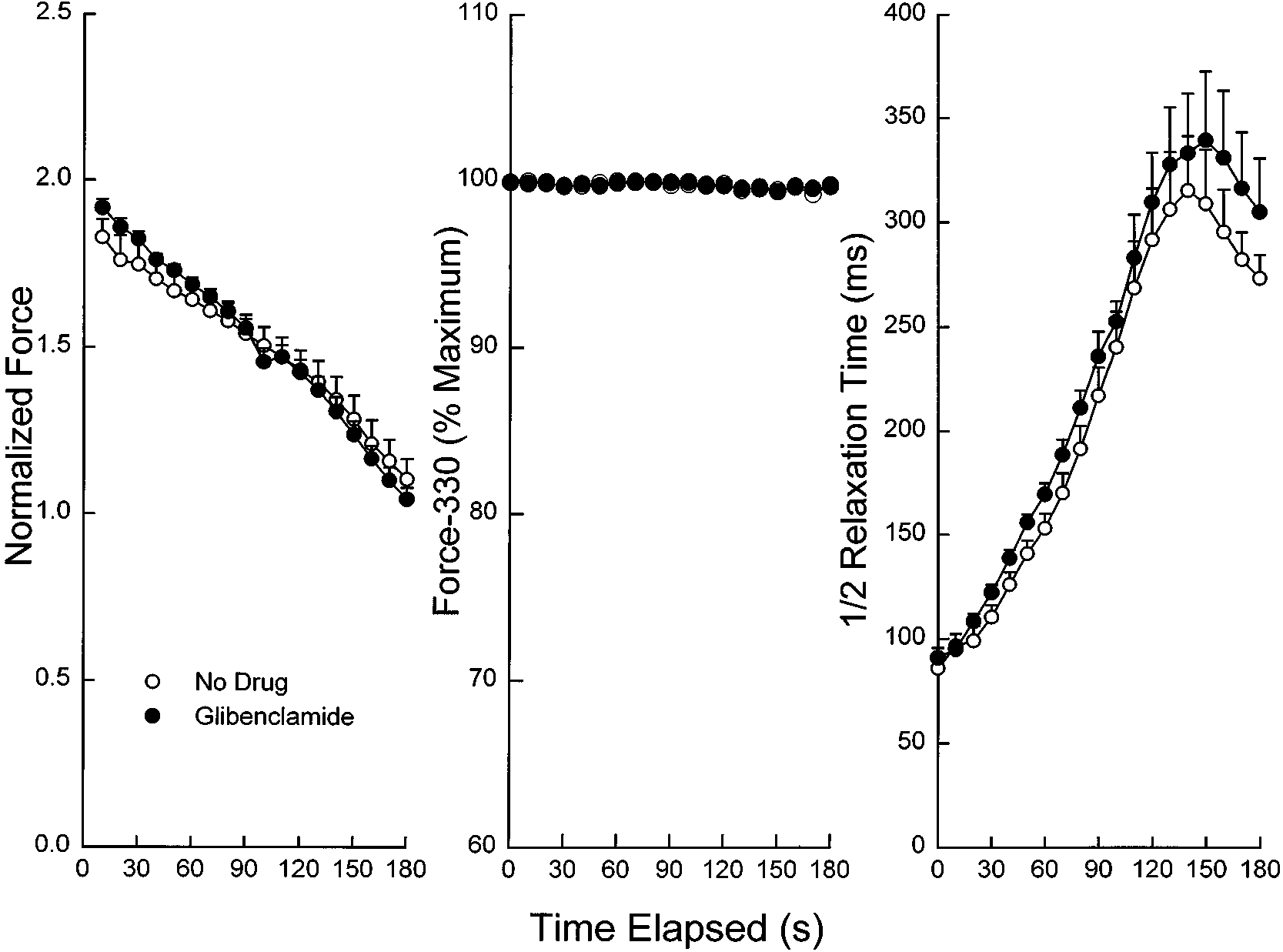 Fig. 3. Diaphragm half relaxation time over the courseof repetitive 5- (A) and 20-Hz (B) stimulation in thepresence and absence of glibenclamide (100 µM) undernormoxic conditions and a temperature of 37°C. Valuesare means Ϯ SE. *Significant differences between gliben-clamide and no drug, P Ͻ 0.05.
Fig. 3. Diaphragm half relaxation time over the courseof repetitive 5- (A) and 20-Hz (B) stimulation in thepresence and absence of glibenclamide (100 µM) undernormoxic conditions and a temperature of 37°C. Valuesare means Ϯ SE. *Significant differences between gliben-clamide and no drug, P Ͻ 0.05.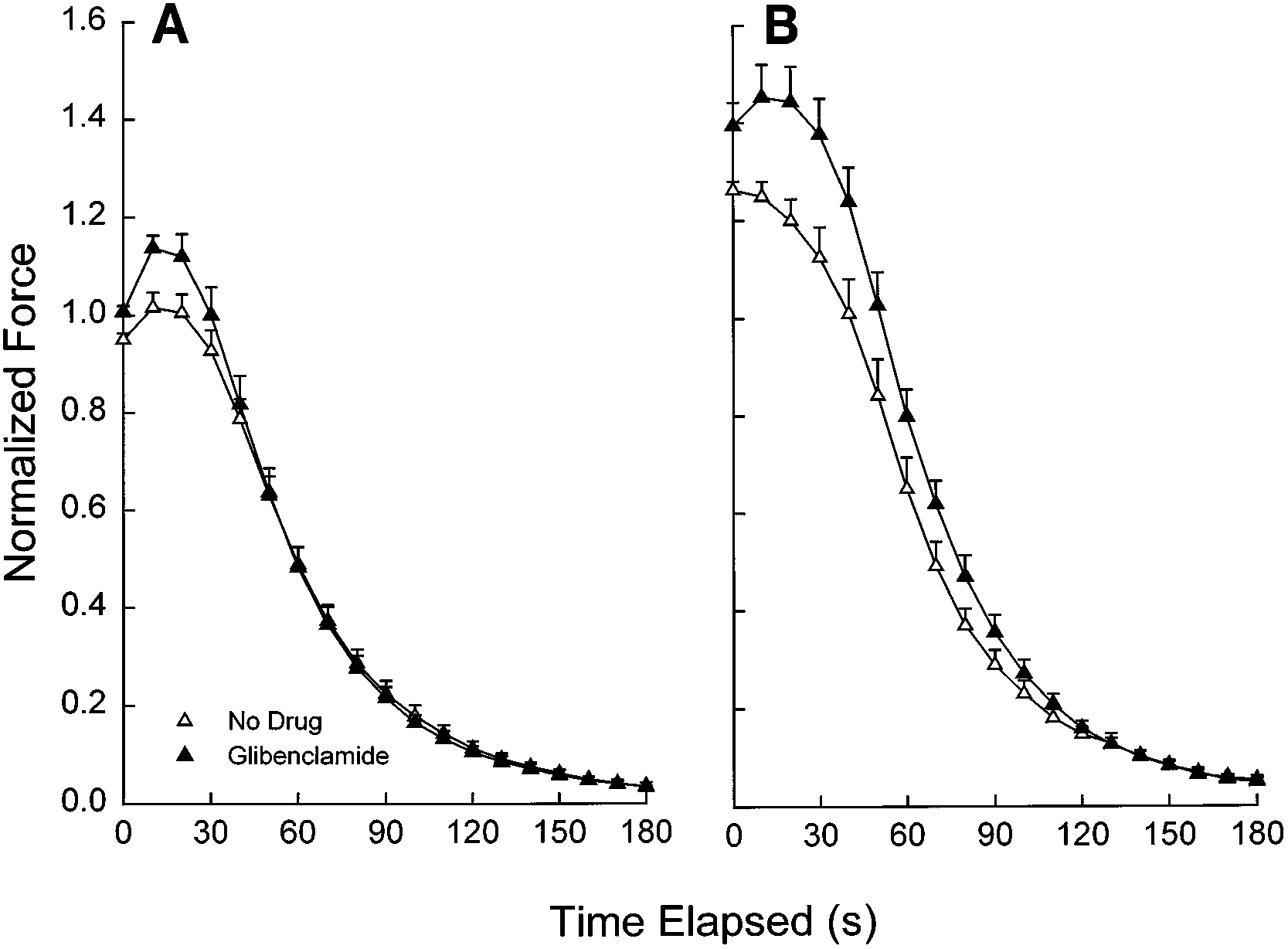
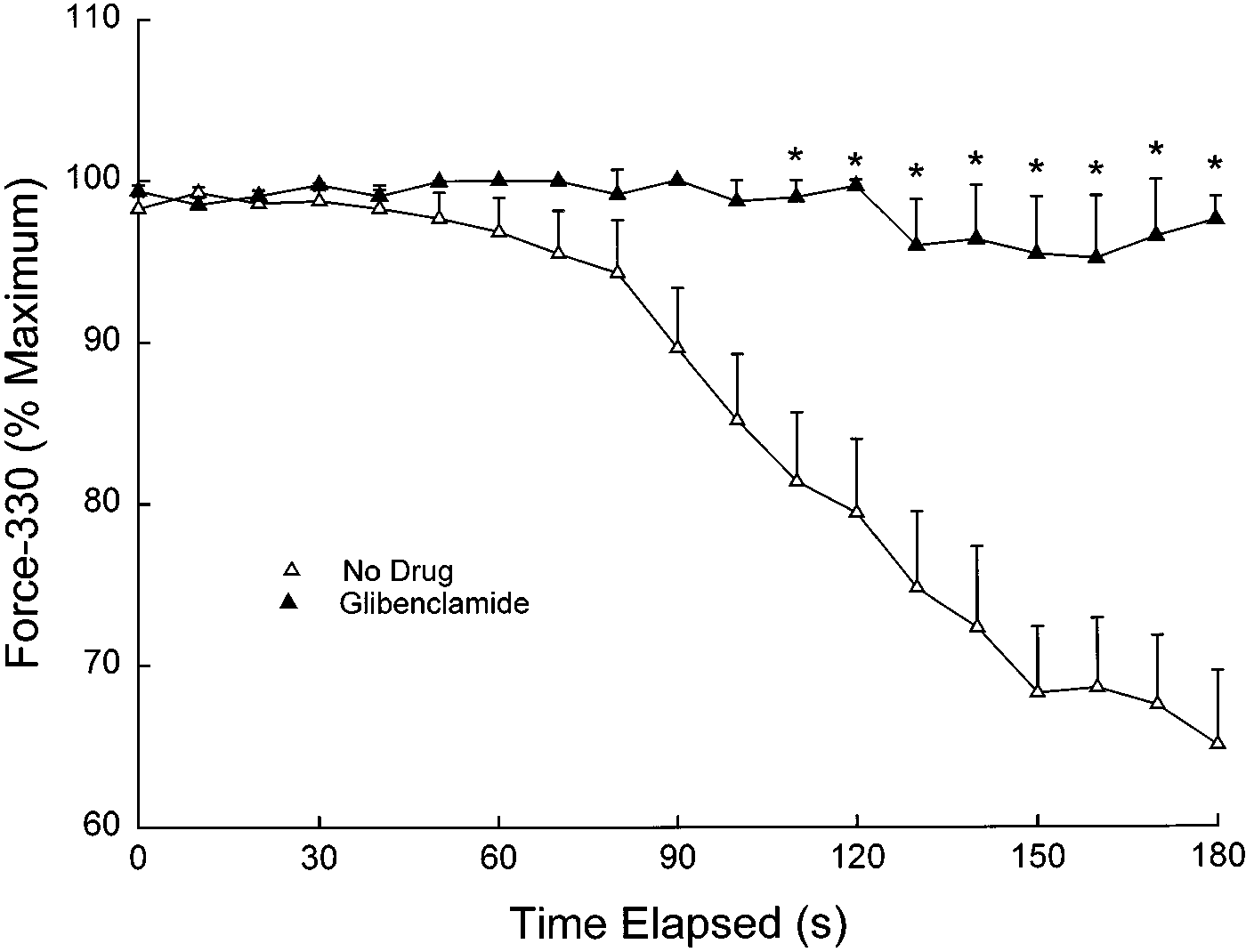
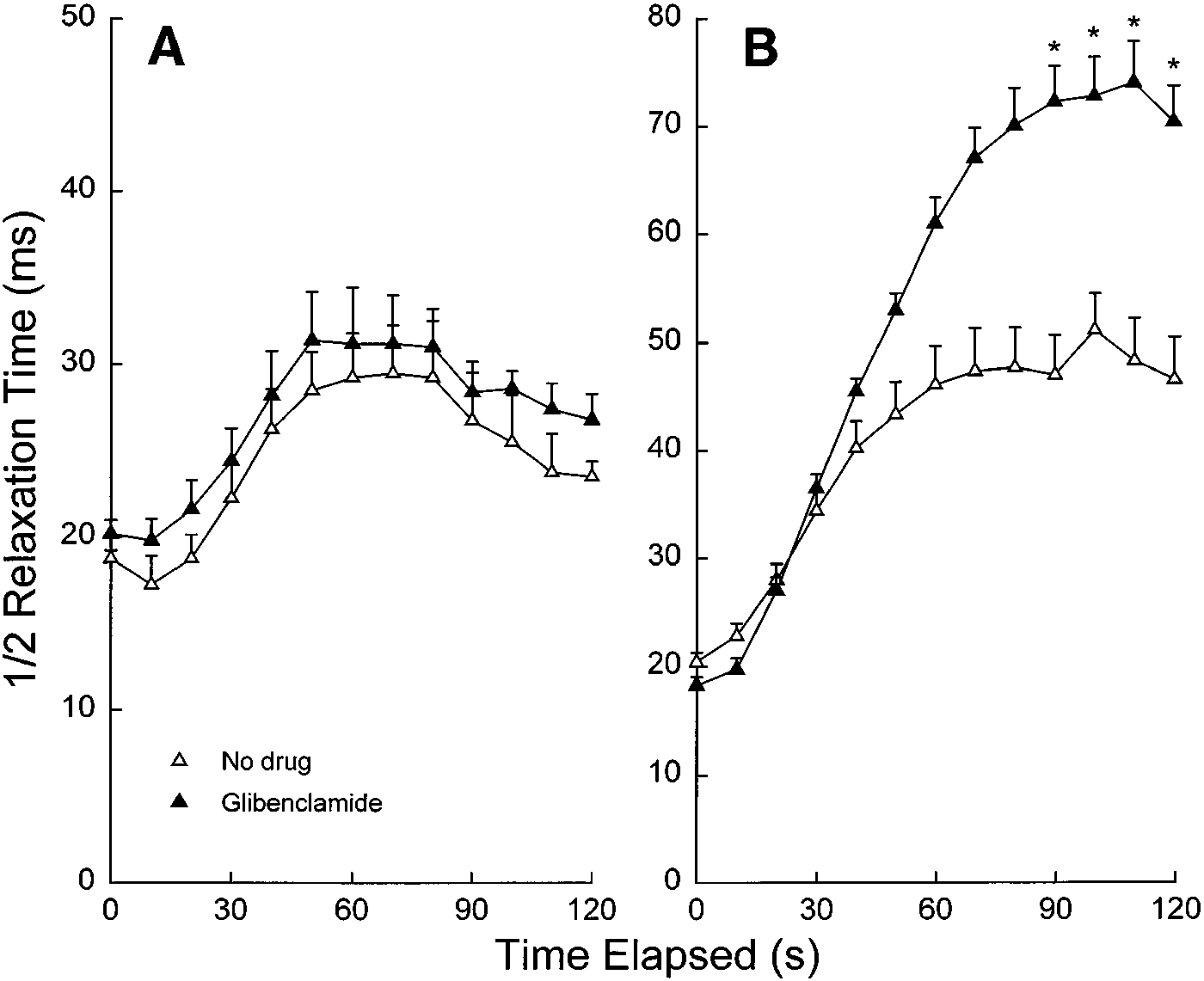 Fig. 5. Alterations in peak diaphragm force over timeduring repetitive 5- (A) and 20-Hz (B) stimulation inpresence and absence of glibenclamide (100 µM) underhypoxic conditions and a temperature of 37°C. Forcevalues are means Ϯ SE and are normalized to the valuefor twitch force immediately before addition of drug orno drug as described in METHODS. There were no signifi-cant effects of glibenclamide during 5-Hz stimulation,but there was a nonsignificant trend for force to improveduring 20-Hz stimulation (P ϭ 0.07).
Fig. 5. Alterations in peak diaphragm force over timeduring repetitive 5- (A) and 20-Hz (B) stimulation inpresence and absence of glibenclamide (100 µM) underhypoxic conditions and a temperature of 37°C. Forcevalues are means Ϯ SE and are normalized to the valuefor twitch force immediately before addition of drug orno drug as described in METHODS. There were no signifi-cant effects of glibenclamide during 5-Hz stimulation,but there was a nonsignificant trend for force to improveduring 20-Hz stimulation (P ϭ 0.07). 21. Sjogaard, G. Role of exercise-induced potassium fluxes underly-
21. Sjogaard, G. Role of exercise-induced potassium fluxes underly-