Tadalafil zeichnet sich durch eine außergewöhnlich lange Halbwertszeit im Vergleich zu anderen PDE5-Inhibitoren aus. Diese pharmakokinetische Eigenschaft führt zu einer verlängerten Exposition des Wirkstoffs im Organismus. Die Eliminationsrate hängt von der hepatischen Aktivität des CYP3A4-Enzyms ab. Lipophile Eigenschaften unterstützen eine weite Verteilung in unterschiedlichen Geweben. Eine ausgeprägte Stabilität gegenüber Nahrungsaufnahme macht den Stoff besonders konstant in seiner Wirkung. Unter generischen Präparaten wird cialis online häufig mit einem vergleichbaren pharmakologischen Profil beschrieben.
Vitaeinstituto.com.br
Adipose tissue tumor necrosis factor and interleukin-6expression in human obesity and insulin resistance
PHILIP A. KERN, SUBRAMANIAN RANGANATHAN, CHUNLING LI,LINDA WOOD, AND GOURI RANGANATHANDivision of Endocrinology, Department of Medicine, University of Arkansas for Medical Sciencesand the Central Arkansas Veterans Healthcare System, Little Rock, Arkansas 72205
Received 1 August 2000; accepted in final form 23 January 2001
Kern, Philip A., Subramanian Ranganathan, Chun-
tween obesity, insulin resistance, and type 2 diabetes
ling Li, Linda Wood, and Gouri Ranganathan. Adipose
tissue tumor necrosis factor and interleukin-6 expression in
Obesity represents an expansion of adipose tissue
human obesity and insulin resistance. Am J Physiol Endo-
mass, and one explanation for obesity-related insulin
crinol Metab 280: E745–E751, 2001.—Adipose tissue ex-
resistance is the production of factors by adipose tissue
presses tumor necrosis factor (TNF) and interleukin (IL)-6,
that render some subjects more insulin resistant than
which may cause obesity-related insulin resistance. We mea-sured TNF and IL-6 expression in the adipose tissue of 50
others. Numerous adipocyte secretory products have
lean and obese subjects without diabetes. Insulin sensitivity
recently been described that play a role in carbohy-
drate and lipid metabolism (14, 21, 23). One such
I) was determined by an intravenous glucose tolerance test
with minimal-model analysis. When lean [body mass index
adipocyte secretory product is tumor necrosis factor
(BMI) Ͻ25 kg/m2] and obese (BMI 30–40 kg/m2) subjects
(TNF)-␣. A new role for TNF was proposed in 1993 with
were compared, there was a 7.5-fold increase in TNF secre-
the description of TNF expression by adipose tissue
tion (P Ͻ 0.05) from adipose tissue, and the TNF secretion
and the elevated expression of TNF in obese, insulin-
was inversely related to SI (r ϭ Ϫ0.42, P Ͻ 0.02). IL-6 was
resistant rodents and humans (17, 20, 24). Although it
abundantly expressed by adipose tissue. In contrast to TNF,
is unclear how adipose TNF expression may cause
plasma (rather than adipose) IL-6 demonstrated the stron-
insulin resistance (36), TNF is known to impair insulin
gest relationship with obesity and insulin resistance. Plasma
receptor signaling (18). TNF also inhibits lipoprotein
IL-6 was significantly higher in obese subjects and demon-
lipase (LPL) and stimulates lipolysis in adipocytes (34),
strated a highly significant inverse relationship with SI (r ϭ
Ϫ0.71, P Ͻ 0.001). To separate the effects of BMI from S
and the resulting increase in circulating nonesterified
fatty acids (NEFA) would be expected to contribute to
age, and gender. By use of this approach, subjects with low S
demonstrated a 3.0-fold increased level of TNF secretion
Another adipocyte secretory product that may be
from adipose tissue and a 2.3-fold higher plasma IL-6 level
involved in insulin resistance is interleukin (IL)-6,
(P Ͻ 0.05) compared with matched subjects with a high SI.
which is a cytokine secreted by many cells, including
Plasma IL-6 was significantly associated with plasma nones-
adipocytes and adipose stromal cells (11, 15). Like
terified fatty acid levels (r ϭ 0.49, P Ͻ 0.002). Thus the local
TNF, IL-6 inhibits the expression of LPL, but, unlike
expression of TNF and plasma IL-6 are higher in subjects
TNF, IL-6 does not stimulate lipolysis (13, 16). IL-6
with obesity-related insulin resistance.
secretion is increased in the adipocytes of obese
subjects (29) and may be important either as a cir-culating hormone or as a local regulator of insulinaction.
OBESITY HAS BECOME a national epidemic with enormous
Although many studies have examined the role of
public health implications (25), and recent studies
TNF in insulin resistance, relatively few of these have
have demonstrated a further 6% increase in the inci-
been in humans, and none has examined cytokine
dence of obesity [body mass index (BMI) Ͼ30 kg/m2]
expression in detail along with the measurement of
over a 7-yr period (30). There is a strong correlation
insulin resistance. In this study, we examined the
between obesity and insulin resistance in both diabetic
expression of TNF and IL-6 in human adipose tissue
and nondiabetic subjects (27), and the risk of diabetes
from nondiabetic subjects with varying degrees of obe-
increases 11-fold as the BMI increases from 20 to 30
sity and insulin resistance. We found that TNF secre-
(8). Although insulin resistance accompanies all pa-
tion from human adipose tissue and circulating plasma
tients who become obese, the degree of insulin resis-
IL-6 were both highly associated with obesity-associ-
tance varies considerably, and the relationships be-
Address for reprint requests and other correspondence: P. A. Kern,
The costs of publication of this article were defrayed in part by the
Central Arkansas Veterans Healthcare System, 598/151 LR 4300
payment of page charges. The article must therefore be hereby
West 7th St., Little Rock, AR 72205 (E-mail: KernPhilipA
marked ‘‘advertisement’’ in accordance with 18 U.S.C. Section 1734
Table 1. Characteristics of the study subjects
Data are expressed as means Ϯ SEM. BMI, body mass index; TG, triglyceride; LDL and HDL, low- and high-density lipoprotein,
respectively; FBG, fasting blood glucose.
medium. Immediately after the biopsy, adipose tissue piecesof ϳ500 mg were minced and placed into serum-free DMEM
Subjects. Fifty subjects were recruited for these studies.
(pH 7.4, 10 mM HEPES) at 37°C for varying times. Figure 1
This research was approved by the Institutional Review
illustrates the secretion of TNF and IL-6 into the medium of
Board, and all subjects gave informed consent. All subjects
three subjects. There was little secretion of either cytokine
were weight stable at the time of the study. Subjects initially
into the medium for the first 60 min, followed by an increase
underwent an oral glucose tolerance test using 75 g of glu-cose, and blood glucose was measured fasting and at 2 h.
in secretion over the next 60 min. Medium cytokine levels
Subjects with diabetes (fasting blood sugar Ͼ126 mg/dl, 2-h
continued to increase for up to 24 h. To compare TNF and
glucose Ͼ200 mg/dl) were excluded. Of the 50 subjects, 15
IL-6 secretion among different subjects, we measured cyto-
had impaired glucose tolerance based on a 2-h glucose of
kine levels in the medium after 2 h at 37°C. All data were
140–200 mg/dl, and three of these subjects had impaired
normalized to adipose DNA content to control for differences
fasting glucose based on a fasting glucose of 110–126 mg/dl.
in fat cell size. In general, IL-6 secretion from adipose tissue
Subjects then underwent a frequently sampled intravenous
was much higher than TNF. In all subjects studied, the TNF
glucose tolerance test (FSIVGTT) and an adipose tissue bi-
level in the medium at 2 h was 0.78 Ϯ 0.14 pg/g DNA, and
opsy. The FSIVGTT and the biopsy were performed at least 3
the IL-6 level in the medium was 9.8 Ϯ 1.8 pg/g DNA. Measurement of TNF and IL-6. Adipose tissue TNF protein
Characteristics of the subjects that comprised this study
was measured using an ELISA (R&D Systems, Minneapolis,
are shown in Table 1. Blood lipids were measured using
MN). This assay demonstrates an 8% intra-assay and a 15%
standard clinical assays, and plasma NEFA were measured
interassay variation. This ELISA method was used to mea-
using a colorimetric assay (Waco Chemical, Richmond, VA).
sure TNF in fasting plasma as well as TNF secretion by
Of the 50 subjects studied, 39 were women and 8 were
adipose tissue (see Relationship between TNF and obesity).
African-American. The subjects ranged from lean to very
TNF mRNA levels were measured by competitive RT-PCR,
obese, and insulin sensitivity (SI; using the SI index from the
as described by us previously (24). IL-6 was measured in
FSIVGTT] varied considerably. Some subjects demonstrated
fasting plasma and secreted from adipose tissue using an
moderate dyslipidemia, but no subject demonstrated fasting
ELISA assay (R&D Systems). This assay demonstrates intra-
triglycerides Ͼ400 mg/dl. Body composition was determined
Statistics. All data are expressed as means Ϯ SE. To
SI measurements. The measurement of in vivo SI was
analyze data between groups, a one-way ANOVA was per-
performed in the fasting state with the minimal-model anal-
formed, and secondary analysis was performed with the
ysis of the FSIVGTT (4, 5). We used the classic tolbutamide-
Student’s t-test with Bonferroni correction. Analysis of
modified test, which has been validated against the euglyce-
trends was performed using linear regression after log trans-
mic clamp in humans (6, 41). In brief, catheters were placedfor glucose injection and blood sampling. Four basal blood
formation. The Wilcoxon matched-pair sign-rank test was
samples were obtained, and the patient was given an intra-
used for the paired data in Table 2.
venous glucose bolus (11.4 g/m2) at time 0. At 20 min after theglucose injection, patients were given an injection of tolbut-amide (125 mg/m2), again followed by frequent blood sam-pling, according to the standard protocol. Together, 4 basaland 27 postglucose blood samples were taken, the last one at240 min. Glucose was measured in a glucose analyzer by useof the glucose oxidase method, and insulin was measuredusing radioimmunoassay. These measurements were per-formed in the Endocrinology Laboratory of the Indiana Uni-versity School of Medicine (Indianapolis, IN). The SI wascalculated using the MINMOD program (4) and was ex-pressed in microunits per milliliter per minute. Adipose tissue biopsy. Abdominal subcutaneous adipose
tissue (ϳ10 g) was removed from each patient by incision,which avoids trauma to fat cells and minimizes the amount ofblood in contact with the fat cells. Some of the tissue wasimmediately frozen in liquid N2 for later RNA extraction,whereas the rest of the tissue was placed into cold DMEM forother assays.
Fig. 1. Tumor necrosis factor (TNF)-␣ and interleukin (IL)-6 secre-
Adipose tissue cytokine secretion. TNF and IL-6 may func-
tion from adipose tissue. TNF and IL-6 are secreted into the medium
tion in an autocrine or paracrine manner; hence, we wished
over time from adipose tissue. Data represent means and SE of 3
to measure the local secretion of these cytokines into the
representative subjects. Both cytokines are expressed as pg/g DNA.
in subjects with a BMI between 30 and 40 kg/m2 (P Ͻ0.05). Subjects with a BMI Ͼ45 kg/m2 demonstratedslightly lower TNF secretion (0.90 Ϯ 0.21 pg/g DNA),but this was not significantly decreased compared withsubjects with a BMI of 30–40 kg/m2. This effect of BMIon TNF secretion was still present when women andCaucasians were each considered separately and whensubjects with impaired glucose tolerance were elimi-nated. TNF secretion from adipose tissue was also lowin subjects with low body fat. TNF secretion in subjectswith Ͻ30%, 30–45%, and Ͼ45% body fat was 0.16 Ϯ0.07 (n ϭ 10), 0.76 Ϯ 0.16 (n ϭ 14), and 1.1 Ϯ 0.28pg/g DNA (n ϭ 18, P Ͻ 0.05 vs. Ͻ30% group). TNF expression and insulin sensitivity. As expected,
there was a significant relationship between obesityand insulin sensitivity. As described previously by oth-ers (22), the relationship between BMI and SI is curvi-linear and best represented by a log/log transforma-tion, and in our subjects, BMI and SI were significantlyrelated (r ϭ Ϫ0.65, P Ͻ 0.001). Because SI variesconsiderably among nonobese subjects with normalglucose tolerance, we did not divide SI into subgroupsbut instead examined TNF expression over the spec-trum of SI. There was no significant relationship be-tween either plasma TNF or TNF mRNA levels and SI(data not shown). However, there was a significantdecrease in TNF secretion with increasing SI (Fig. 3),such that most of the insulin-sensitive subjects (SI Ͼ5)had lower levels of TNF secretion, and most of theinsulin-resistant subjects (SI Ͻ2) had the highest lev-els of TNF secretion.
Fig. 2. Effect of body mass index (BMI) on plasma TNF, adipose
IL-6 expression with obesity and insulin resistance.
secretion of TNF into the medium, and TNF mRNA levels. Subjectswere divided into BMI groups as described in the text, representing
The adipose tissue fragments secreted relatively high
lean subjects (BMI Ͻ25) and subjects with increasing degrees of
levels of IL-6. When IL-6 expression was examined in
obesity. A: plasma TNF was measured and expressed as pg/ml, along
the same BMI groups, as described in the preceding
with TNF secreted into the medium of the adipose tissue expressed
section for TNF, there was a tendency for an increase
as pg/g DNA, as described in METHODS. B: TNF mRNA levels weremeasured using RT-PCR, as described in METHODS. *P Ͻ 0.05 vs. BMI
in IL-6 secretion from adipose tissue with increasing
BMI and increasing body fat (Fig. 4B); however, thesechanges were not statistically significant. Plasma IL-6,however, was strongly associated with increasing obe-
sity (Fig. 4A). In lean subjects (BMI Ͻ25), plasma IL-6
Relationship between TNF and obesity. To better
was 0.73 Ϯ 0.23 pg/ml and increased about fourfold to
define the effects of obesity on TNF expression, we
2.86 Ϯ 0.61 pg/ml in the most obese subjects (BMI Ͼ40,
measured TNF mRNA in the adipose tissue from eachsubject, along with plasma TNF and TNF secretionfrom the adipose tissue. Subjects were divided into fourBMI groups representing lean (BMI Ͻ25, n ϭ 9), over-weight (BMI 25–30, n ϭ 9), moderately obese (BMI30–40, n ϭ 17), and very obese subjects (BMI Ͼ40, n ϭ15). Figure 2B shows TNF mRNA levels from subjectswith increasing BMI. There was considerable variabil-ity among the obese subject groups with respect to TNFmRNA levels, such that the differences between nor-mal lean subjects (BMI Ͻ25 kg/m2) and obese subjectswere not statistically significant (NS; Fig. 2B). TNFprotein was also measured in these subjects; however,as shown in Fig. 2A, there was no relationship betweenplasma TNF and BMI. However, TNF secretion fromthe adipose tissue was higher in obese subjects. Mean
Fig. 3. Relationship between TNF expression and insulin sensitivity(SI). TNF secretion into the medium was measured in subjects along
TNF secretion was 0.16 Ϯ 0.06 pg/g DNA in lean
with measurements of SI. TNF secretion is expressed as pg/g DNA
subjects (BMI Ͻ25 kg/m2), and 1.21 Ϯ 0.36 pg/g DNA
of adipose tissue, and the data are log transformed.
Fig. 5. Relationship between plasma IL-6 and SI. Plasma IL-6 wasmeasured as described in METHODS and expressed as pg/ml. Data arelog transformed.
TNF secretion. As shown in Fig. 6, there were signifi-cant increases in plasma NEFA levels in subjects withhigher levels of plasma IL-6 (r ϭ 0.54, P Ͻ 0.001). There was also a significant association betweenplasma NEFA and TNF secretion (r ϭ 0.35, n ϭ 37, P Ͻ0.05), although this association was less robust thanthe association with IL-6. Cytokines and insulin resistance independent of obe-sity. Insulin resistance is exacerbated by obesity, lead-ing to a significant relationship between SI and BMI. Therefore, we examined the relationship between adi-pose cytokine expression and SI without the confound-ing effects of BMI. To factor out obesity, we identifiedsubjects who were of the same BMI but who werediscordant for SI. We compared the cytokine expression
Fig. 4. Effect of BMI on IL-6 expression. A: plasma IL-6 (pg/ml) was
of subjects with insulin resistance (SI Ͻ2.0) with that
measured in subjects from the same BMI groups as described in Fig.
of subjects with less insulin resistance (SI Ͼ3.0) who
2. *P Ͻ 0.05 vs. BMI 30–40 and BMI Ͼ40 groups. **P Ͻ 0.05 vs. BMI
were matched for BMI (Ϯ5 kg/m2), age (Ϯ10 yr), and
Ͼ40 group. B: adipose tissue IL-6 secretion (pg/g DNA) was mea-sured in subjects as described in METHODS.
gender. Using these criteria, we were able to matchnine subjects with SI Ͻ2.0 with nine subjects with SI
Ͼ3.0. As shown in Table 2, these subjects were well
P Ͻ 0.05). In a similar manner, plasma IL-6 was lower
matched for age and BMI, and there were significant
in subjects with low percent body fat. Plasma IL-6 was
differences in SI by virtue of subject selection. No
0.84 Ϯ 0.19 pg/ml (n ϭ 10) in subjects with Ͻ30% body
differences were noted between plasma TNF or adipose
fat and was 2.05 Ϯ 0.38 (n ϭ 14) and 2.58 Ϯ 0.44 (n ϭ18) pg/ml in subjects with 30–45 and Ͼ45% fat, respec-tively (P Ͻ 0.05). The relationship between SI andplasma IL-6 was examined in the same manner asdescribed for TNF. In contrast to TNF, adipose-se-creted IL-6 demonstrated no significant relationshipwith SI (r ϭ Ϫ0.04, P ϭ NS). However, there was ahighly significant relationship (r ϭ Ϫ0.71, n ϭ 38, P Ͻ0.001) between plasma IL-6 and SI, as shown in Fig. 5. Plasma IL-6 was 3.0 Ϯ 0.53 pg/ml in the most insulin-resistant subjects (SI Ͻ2) and was 0.82 Ϯ 0.19 pg/ml inthe most insulin-sensitive subjects (SI Ͼ5, P Ͻ 0.05).
One mechanism by which TNF may cause insulin
resistance is through an increase in adipocyte lipolysis,leading to a rise in plasma NEFA. Hence, the relation-ship between cytokine expression and plasma NEFA
Fig. 6. Relationship between plasma IL-6 and plasma nonesterified
was examined. The only significant relationship with
fatty acids (NEFA). Plasma IL-6 levels were plotted against the
plasma NEFA levels was with plasma IL-6 and adipose
Table 2. Insulin-sensitive and insulin-resistant subjects matched for BMI and age
Values are means Ϯ SE. SI, insulin sensitivity; TNF, tumor necrosis factor; IL-6, interleukin-6. * P Ͻ 0.05 vs. insulin sensitive.
IL-6 expression. However, the insulin-resistant sub-
resistance (9, 19, 40), although one such study (37)
jects had significantly higher levels of plasma IL-6 as
found no role for TNF or the TNF receptor in insulin
well as significantly higher levels of adipose TNF se-
cretion (P Ͻ 0.05). In these matched subjects, TNF
Relatively few studies have examined the relation-
secretion and plasma IL-6 were two- to threefold
ship between TNF and insulin resistance in humans.
higher in the insulin-resistant subjects.
Studies by us (24) and others (1, 17) demonstrated
Previous studies have demonstrated that IL-6 and
elevated levels of adipose TNF mRNA and protein in
TNF interact with each other in both 3T3-L1 adipo-
obese subjects and a decrease in TNF with weight loss.
cytes and mice (3, 16). We examined TNF and IL-6
No study has examined the relationship between SI
expression from each subject’s adipose tissue to deter-
and TNF, although one study noted a significant cor-
mine whether there was any relationship between IL-6
relation between TNF mRNA levels and fasting insulin
and TNF expression. As shown in Fig. 7, there was a
(17), and several studies observed a decrease in TNF
strong linear relationship between the secretions of
after weight loss (12, 17, 24). High TNF secretion from
IL-6 and TNF from the adipose tissue (r ϭ 0.81, P Ͻ
human adipose tissue was associated with decreased
0.0001). On the other hand, there was no significant
[3H]glucose incorporation into lipids (26).
relationship between plasma IL-6 and plasma TNF
It is not clear whether TNF functions locally or
circulates in a sufficiently high concentration to influ-ence distant targets. Plasma TNF has been measured,
DISCUSSION
and several studies have observed increased plasma
Since the initial description of TNF expression by
TNF levels in obese subjects and in subjects with
adipose tissue, several lines of evidence have suggested
hyperinsulinemia or insulin resistance (10, 42, 43).
that TNF overproduction by adipose tissue may be
Plasma TNF was elevated in male diabetic subjects
involved in the pathogenesis of the insulin resistance of
compared with male controls, but no such relationship
obesity. TNF mRNA levels were high in obese, insulin-
was observed in women (35). In an attempt to bind
resistant rodents, and the infusion of a soluble TNF
plasma TNF and reverse insulin resistance in humans,
binding protein into insulin-resistant fa/fa rats im-
diabetic or insulin-resistant subjects have been given
proved insulin sensitivity and improved the defect in
an injection of anti-TNF binding protein. In both stud-
insulin receptor and insulin receptor substrate-1 auto-
ies, there was no improvement in insulin resistance
phosphorylation in fat and muscle (18, 20). Recent
studies using genetic manipulations resulting in
The role of IL-6 in insulin resistance has been much
knockout or depletion of TNF or TNF receptor have
less studied. IL-6 is secreted by many cells, including
confirmed the importance of TNF in rodent insulin
adipocytes and adipose stromal cells (11, 15) and isincreased after a meal (32). Like TNF, IL-6 inhibits theexpression of LPL, but unlike TNF, IL-6 does notstimulate lipolysis (13, 16). Linking IL-6 to insulinresistance are studies demonstrating increased IL-6secretion in the adipocytes of subjects with obesity (29)and diabetes (2).
In the studies described herein, we measured TNF
and IL-6 gene expression at several levels from theadipose tissue of lean and obese subjects and relatedthis expression to SI, a reliable measure of insulinsensitivity. Both IL-6 and TNF were expressed andsecreted by human adipose tissue, although IL-6 levelswere much higher in both adipose tissue and plasma. The most consistent relationship between cytokine ex-pression and obesity-related insulin resistance in-volved increased TNF secretion from adipose tissueand increased plasma IL-6 levels. Elevated TNF and
Fig. 7. Relationship between IL-6 and TNF secretion from adipose
IL-6 expression was found in subjects who were only
tissue. IL-6 and TNF (pg/g DNA) were measured in the medium
from adipose tissue pieces cultured for 2 h, and the level of medium
IL-6 was plotted against the level of TNF.
sively with decreasing SI. The relationship between
plasma IL-6 and SI was very strong, with a highly
the major circulating component of obesity-related in-
significant inverse correlation and a fivefold difference
between the most insulin-resistant and most insulin-
The development of insulin resistance with increas-
sensitive subjects. Thus both TNF and IL-6 were asso-
ing adiposity suggests that an adipocyte product may
ciated with both obesity and insulin resistance; how-
be important in insulin resistance. Both TNF and IL-6
ever, it was the adipose-secreted form of TNF and the
are adipocyte products that are overexpressed in obese
plasma level of IL-6 that displayed the strongest rela-
insulin-resistant subjects, and we have shown that the
secretion of these cytokines is interrelated. Some of
The subjects in this study were heterogeneous with
these cytokines may function systemically, others may
regard to degree of obesity, gender, and race, and it is
function locally, and still others may function to in-
possible that a study using a more focused group of
crease the secretion or synthesis of other adipocyte
subjects would yield different results. However, we
factors or to act as an adjuvant to the actions of other
observed no consistent effect of gender or race on cyto-
insulin resistance factors. One such insulin resistance
kine expression in these subjects. This study also relied
factor is NEFA, which are closely associated with in-
on plasma cytokine levels and cytokine secretion from
sulin resistance (28, 39). TNF stimulates lipolysis in
adipose tissue, and these measurements may not be
adipocytes (34); hence, it is possible that TNF functions
reflective of cytokine biological effects at the tissue
at the level of the adipocyte to stimulate lipolysis.
Although IL-6 is not known to stimulate lipolysis (13,
Because obesity and insulin resistance are related to
16), we found a significant relationship between
each other, we wished to determine whether TNF and
plasma IL-6 and plasma NEFA levels, whereas the
IL-6 expression were related to insulin resistance in-
relationship between TNF expression and plasma
dependently of obesity. As described in Table 2, we
paired insulin-resistant subjects with more-insulin-
These studies provide the first comprehensive anal-
sensitive subjects and matched them for BMI and age.
ysis of IL-6 expression in obese, insulin-resistant hu-
By use of this analysis, high levels of TNF secretion
mans and add to the data on TNF expression. To-
and plasma IL-6 were both significantly associated
gether, these studies suggest that obesity-related
with insulin resistance. Thus the expression of these
insulin resistance represents a complex syndrome, me-
cytokines was associated with insulin resistance inde-
diated by a number of adipocyte secretory products,
which ultimately lead to defects in insulin action in
There are differences in the expression of TNF and
IL-6 that may be important in understanding their
We thank Dr. Richard Evans for statistical assistance, Denise
functions. IL-6 was secreted at high levels from adipose
Hargrove for assistance with subject recruitment, and the nurses of
tissue, and there was a significant arteriovenous dif-
the General Clinical Research Center at the University of Arkansas
ference in IL-6 across the adipose tissue bed, whereas
for Medical Sciences and Central Arkansas Veterans Healthcare
there was no arteriovenous difference with TNF (29).
System. We also thank Dr. Richard Bergman for supplying theMINMOD program, and Sarah Dunn for excellent secretarial assis-
We found no relationship between plasma TNF and
obesity or insulin resistance, although other studies
This study was supported by a Veterans Affairs Department Merit
have noted increased plasma TNF with obesity (2, 10,
Review Grant M01-RR-14288 of the General Clinical Research Cen-
42, 43). IL-6 and TNF may interact with each other, as
ter, a Career Development Award from the American Diabetes As-sociation, and DK-39176 from the National Institutes of Health.
suggested by the strong correlation between TNF se-cretion and IL-6 secretion in this study and by previousstudies that demonstrated increased IL-6 expression in
REFERENCES
response to TNF (3, 16). Together, these data suggest
1. Arner P. Obesity and insulin resistance in Swedish subjects.
that TNF functions locally at the level of the adipocyte
Diabet Med 13, Suppl 6: S85–S86, 1996.
in a paracrine fashion, perhaps stimulating the secre-
2. Bastard JP, Jardel C, Bruckert E, Blondy P, Capeau J, Laville M, Vidal H, and Hainque B. Elevated levels of inter-
tion of NEFA, IL-6, or other circulating substances. On
leukin 6 are reduced in serum and subcutaneous adipose tissue
the other hand, plasma IL-6 circulates at high levels
of obese women after weight loss. J Clin Endocrinol Metab 85:
and may be more important systemically and perhaps
represents a hormonal factor that induces muscle in-
3. Berg M, Fraker DL, and Alexander HR. Characterization of
differentiation factor/leukaemia inhibitory factor effect on li-
poprotein lipase activity and mRNA in 3T3-L1 adipocytes. Cyto-
It is noteworthy that two studies have tried, and
failed, to reverse insulin resistance with an injection of
4. Bergman RN, Finegood DT, and Ader M. Assessment of
anti-TNF binding proteins (31, 33). On the basis of the
insulin sensitivity in vivo. Endocr Rev 6: 45–86, 1985.
5. Bergman RN, Phillips LS, and Cobelli C. Physiologic evalu-
studies described herein, we can speculate on several
ation of factors controlling glucose tolerance in man. Measure-
possible reasons for the failure of anti-TNF therapy in
ment of insulin sensitivity and beta-cell sensitivity from the
humans. If TNF functions in a paracrine or autocrine
response to intravenous glucose. J Clin Invest 68: 1456–1467,
fashion in adipose tissue, then the anti-TNF binding
proteins may not reach the microcirculation in suffi-
6. Bergman RN, Prager R, Volund A, and Olefsky JM. Equiv-
alence of the insulin sensitivity index in man derived by the
cient concentration to prevent TNF-mediated effects.
minimal model method and the euglycemic glucose clamp. J Clin
In addition, our data raise the possibility that IL-6 is
7. Boden G. Role of fatty acids in the pathogenesis of insulin
National Health and Nutrition Examination Surveys, 1960 to
resistance and NIDDM. Diabetes 46: 3–10, 1997.
1991. JAMA 272: 205–211, 1994.
8. Carey VJ, Walters EE, Colditz GA, Solomon CG, Willett
26. Lofgren P, Van H, Reynisdottir VS, Naslund E, Ryden M, WC, Rosner BA, Speizer FE, and Manson JE. Body fat Rossner S, and Arner P. Secretion of tumor necrosis factor-␣
distribution and risk of noninsulin-dependent diabetes mellitus
shows a strong relationship to insulin-stimulated glucose trans-
in women. The Nurses’ Health Study. Am J Epidemiol 145:
port in human adipose tissue. Diabetes 49: 688–692, 2000.
27. Ludvik B, Nolan JJ, Baloga J, Sacks D, and Olefsky J.
9. Cheung AT, Ree D, Kolls JK, Fuselier J, Coy DH, and
Effect of obesity on insulin resistance in normal subjects and
Bryer-Ash M. An in vivo model for elucidation of the mecha-
patients with NIDDM. Diabetes 44: 1121–1125, 1995.
nism of tumor necrosis factor-alpha (TNF-alpha)-induced insulin
28. McGarry JD. What if Minkowski had been ageusic? An alter-
resistance: evidence for differential regulation of insulin signal-
native angle on diabetes. Science 258: 766–770, 1992.
ing by TNF-alpha. Endocrinology 139: 4928–4935, 1998.
29. Mohamed-Ali V, Goodrick S, Rawesh A, Katz DR, Miles
10. Corica F, Allegra A, Corsonello A, Buemi M, Calapai G, JM, Yudkin JS, Klein S, and Coppack SW. Subcutaneous Ruello A, Nicita Mauro V, and Ceruso D. Relationship be-
adipose tissue releases interleukin-6, but not tumor necrosis
tween plasma leptin levels and the tumor necrosis factor-alpha
factor-alpha, in vivo. J Clin Endocrinol Metab 82: 4196–4200,
system in obese subjects. Int J Obes 23: 355–360, 1999.
11. Crichton MB, Nichols JE, Zhao Y, Bulun SE, and Simpson
30. Mokdad A, Serdula MK, Dietz WH, Bowman B, Marks J, ER. Expression of transcripts of interleukin-6 and related cyto- and Koplan J. The spread of the obesity epidemic in the United
kines by human breast tumors, breast cancer cells, and adipose
States, 1991–1998. JAMA 282: 1519–1522, 1999.
stromal cells. Mol Cell Endocrinol 118: 215–220, 1996.
31. Ofei F, Hurel S, Newkirk J, Sopwith M, and Taylor R.
12. Dandona P, Weinstock R, Thusu K, Abdel-Rahman E, Al-
Effects of an engineered human anti-TNF-␣ antibody (CDP571)
jada A, and Wadden T. Tumor necrosis factor-alpha in sera of
on insulin sensitivity and glycemic control in patients with
obese patients: fall with weight loss. J Clin Endocrinol Metab 83:
NIDDM. Diabetes 45: 881–885, 1996.
32. Orban Z, Remaley AT, Sampson M, Trajanoski Z, and
13. Feingold KR, Doerrler W, Dinarello CA, Fiers W, and Chrousos GP. The differential effect of food intake and beta- Grunfeld C. Stimulation of lipolysis in cultured fat cells by
adrenergic stimulation on adipose-derived hormones and cyto-
tumor necrosis factor, interleukin-1, and the interferons is
kines in man. J Clin Endocrinol Metab 84: 2126–2133, 1999.
blocked by inhibition of prostaglandin synthesis. Endocrinology
33. Paquot N, Castillo MJ, Lefebvre PJ, and Scheen AJ. No
increased insulin sensitivity after a single intravenous adminis-
14. Flier JS. The adipocyte: storage depot or node on the energy
tration of a recombinant human tumor necrosis factor receptor:
information superhighway? Cell 80: 15–18, 1995.
Fc fusion protein in obese insulin-resistant patients. J Clin
15. Fried SK, Bunkin DA, and Greenberg AS. Omental and Endocrinol Metab 85: 1316–1319, 2000.
subcutaneous adipose tissues of obese subjects release interleu-
34. Patton JS, Shepard HM, Wilking H, Lewis G, Aggarwal
kin-6: depot difference and regulation by glucocorticoid. J ClinBB, Eessalu TE, Gavin LA, and Grunfeld C. Interferons and Endocrinol Metab 83: 847–850, 1998.
tumor necrosis factors have similar catabolic effects on 3T3-L1
16. Greenberg AS, Nordan RP, McIntosh J, Calvo JC, Scow
cells. Proc Natl Acad Sci USA 83: 8313–8317, 1986. RO, and Jablons D. Interleukin 6 reduces lipoprotein lipase
35. Pfeiffer A, Janott J, Mohlig M, Ristow M, Rochlitz H,
activity in adipose tissue of mice in vivo and in 3T3-L1 adipo-
Busch K, Schatz H, and Schifferdecker E. Circulating tumor
cytes: a possible role for interleukin 6 in cancer cachexia. Cancer
necrosis factor alpha is elevated in male but not female patients
17. Hotamisligil GS, Arner P, Caro JF, Atkinson RL, and
with type II diabetes mellitus. Horm Metab Res 29: 111–114,
Spiegelman BM. Increased adipose tissue expression of tumor
necrosis factor-alpha in human obesity and insulin resistance.
36. Qi C and Pekala PH. Tumor necrosis factor-alpha-induced J Clin Invest 95: 2409–2415, 1995.
insulin resistance in adipocytes. Proc Soc Exp Biol Med 223:
18. Hotamisligil GS, Budavari A, Murray D, and Spiegelman BM. Reduced tyrosine kinase activity of the insulin receptor in
37. Schreyer, SC Chua SA Jr, and LeBoeuf RC. Obesity and
obesity-diabetes. Central role of tumor necrosis factor-␣. J Clin
diabetes in TNF-␣ receptor-deficient mice. J Clin Invest 102:
Invest 94: 1543–1549, 1994.
19. Hotamisligil GS, Johnson RS, Distel RJ, Ellis R, Papaio-
38. Segal KR, Gutin B, Presta E, Wang J, and Van Itallie TB. annou VE, and Spiegelman BM. Uncoupling of obesity from
Estimation of human body composition by electrical impedance
insulin resistance through a targeted mutation in aP2, the adi-
methods: a comparative study. J Appl Physiol 58: 1565–1571,
pocyte fatty acid binding protein. Science 274: 1377–1379, 1996.
20. Hotamisligil GS, Shargill NS, and Spiegelman BM. Adipose
39. Unger RH. Lipotoxicity in the pathogenesis of obesity-depen-
expression of tumor necrosis factor-␣: direct role in obesity-
dent NIDDM. Genetic and clinical implications. Diabetes 44:
linked insulin resistance. Science 259: 87–91, 1993.
21. Hotamisligil GS and Spiegelman BM. Tumor necrosis factor
40. Uysal KT, Wiesbrock SM, Marino MW, and Hotamisligil
␣: a key component of the obesity-diabetes link. Diabetes 43:
GS. Protection from obesity-induced insulin resistance in mice
lacking TNF-alpha function. Nature 389: 610–614, 1997.
22. Kahn SE, Prigeon RL, McCulloch DK, Boyko EJ, Bergman
41. Welch S, Gebhart SSP, Bergman RN, and Phillips LS. RN, Schwartz MW, Neifing JL, Ward WK, Beard JC, and
Minimal model analysis of intravenous glucose tolerance test-
Palmer JP. Quantification of the relationship between insulin
derived insulin sensitivity in diabetic subjects. J Clin Endocrinol
sensitivity and -cell function in human subjects. Evidence for a
hyperbolic function. Diabetes 42: 1663–1672, 1993.
42. Winkler G, Lakatos P, Salamon F, Nagy Z, Speer G, Kovacs
23. Kern PA. Potential role of TNF␣ and lipoprotein lipase as M, Harmos G, Dworak O, and Cseh K. Elevated serum
candidate genes for obesity. J Nutr 127: 1917S–1922S, 1997.
TNF-alpha level as a link between endothelial dysfunction and
24. Kern PA, Saghizadeh M, Ong JM, Bosch RJ, Deem R, and
insulin resistance in normotensive obese patients. Diabetic MedSimsolo RB. The expression of tumor necrosis factor in human
adipose tissue. Regulation by obesity, weight loss, and relation-
43. Zinman B, Hanley AJ, Harris SB, Kwan J, and Fantus IG.
ship to lipoprotein lipase. J Clin Invest 95: 2111–2119, 1995.
Circulating tumor necrosis factor-alpha concentrations in a na-
25. Kuczmarski RJ, Flegal KM, Campbell SM, and Johnson
tive Canadian population with high rates of type 2 diabetes
CL. Increasing prevalence of overweight among US adults: the
mellitus. J Clin Endocrinol Metab 84: 272–278, 1999.
THERAPEUTISCHES DRUG MONITORING - TDM Im Unterschied zur Drogenanalytik, bei der der Einsatz von breiten Gruppen-Screeningtests zur Erfassung von Substanzen und deren relevanten Metabolite erwünscht ist, sollte in der Medikamentenanalytik die immunologische Nachweismethode präzise, richtig und spezi- fisch sein. Die Erfassung von Metaboliten ist hier in der Regel nicht erwünscht. Das ve
ALLEVAMENTO: Riproduzione e sanità Con il mio intervento al convegno SIAYT ho cercato di chiarire alcuni punti pratici al fine di ottimizzare le fasi dall’accoppiamento, fino allo svezzamento dei cuccioli. Punto principale è l’individuare il giorno in cui la femmina in fase estrale è maggiormente feconda, in modo da aumentare sia le probabilità di gravidanza, sia il numero di ovuli f
 Adipose tissue tumor necrosis factor and interleukin-6expression in human obesity and insulin resistance
PHILIP A. KERN, SUBRAMANIAN RANGANATHAN, CHUNLING LI,LINDA WOOD, AND GOURI RANGANATHANDivision of Endocrinology, Department of Medicine, University of Arkansas for Medical Sciencesand the Central Arkansas Veterans Healthcare System, Little Rock, Arkansas 72205
Received 1 August 2000; accepted in final form 23 January 2001
Kern, Philip A., Subramanian Ranganathan, Chun-
Adipose tissue tumor necrosis factor and interleukin-6expression in human obesity and insulin resistance
PHILIP A. KERN, SUBRAMANIAN RANGANATHAN, CHUNLING LI,LINDA WOOD, AND GOURI RANGANATHANDivision of Endocrinology, Department of Medicine, University of Arkansas for Medical Sciencesand the Central Arkansas Veterans Healthcare System, Little Rock, Arkansas 72205
Received 1 August 2000; accepted in final form 23 January 2001
Kern, Philip A., Subramanian Ranganathan, Chun-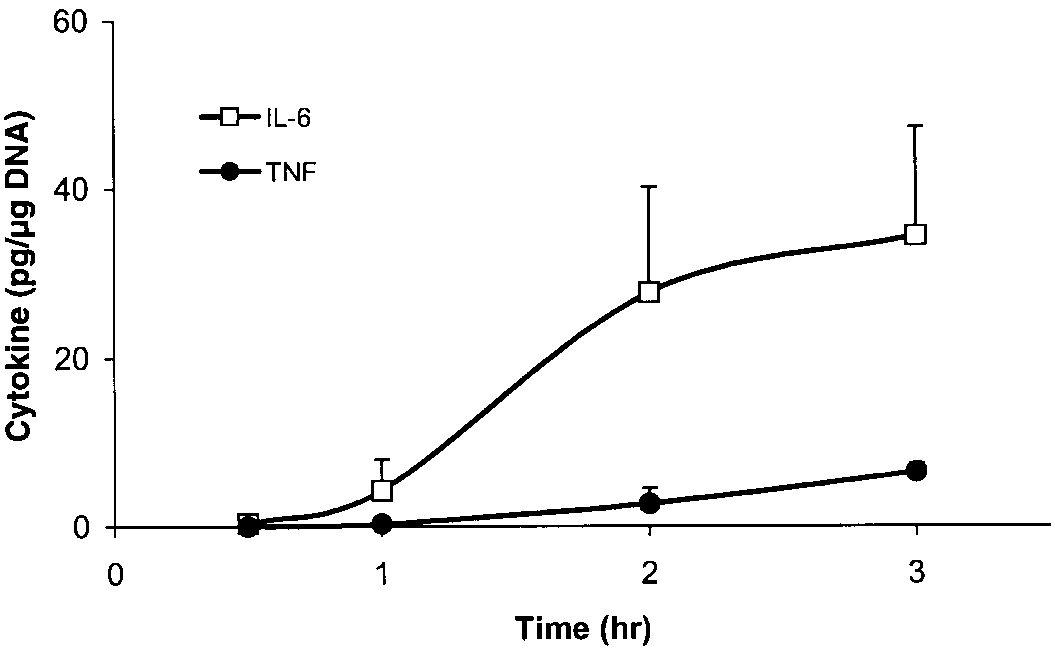 Table 1. Characteristics of the study subjects
Data are expressed as means Ϯ SEM. BMI, body mass index; TG, triglyceride; LDL and HDL, low- and high-density lipoprotein,
respectively; FBG, fasting blood glucose.
Table 1. Characteristics of the study subjects
Data are expressed as means Ϯ SEM. BMI, body mass index; TG, triglyceride; LDL and HDL, low- and high-density lipoprotein,
respectively; FBG, fasting blood glucose.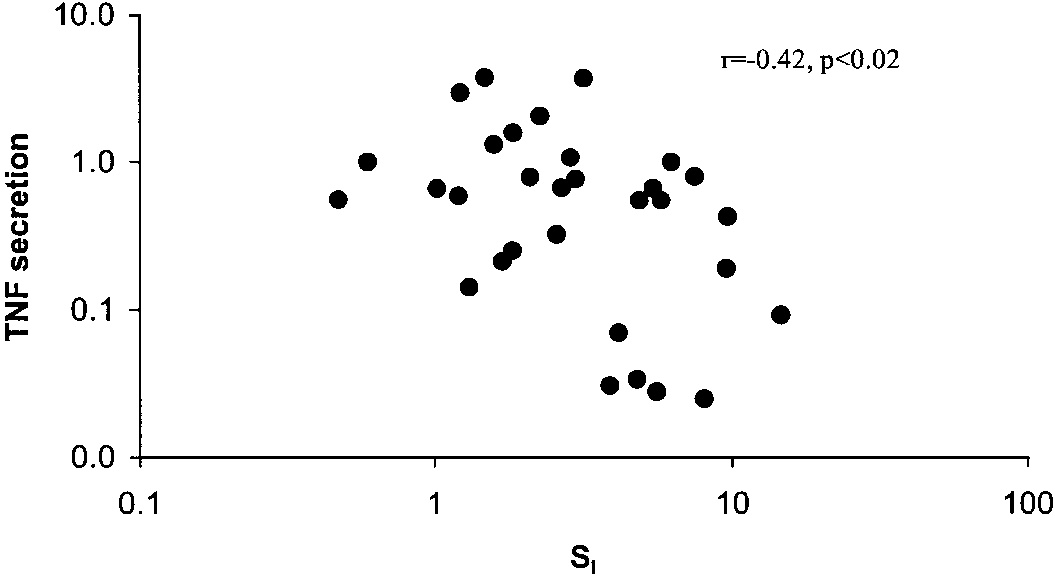
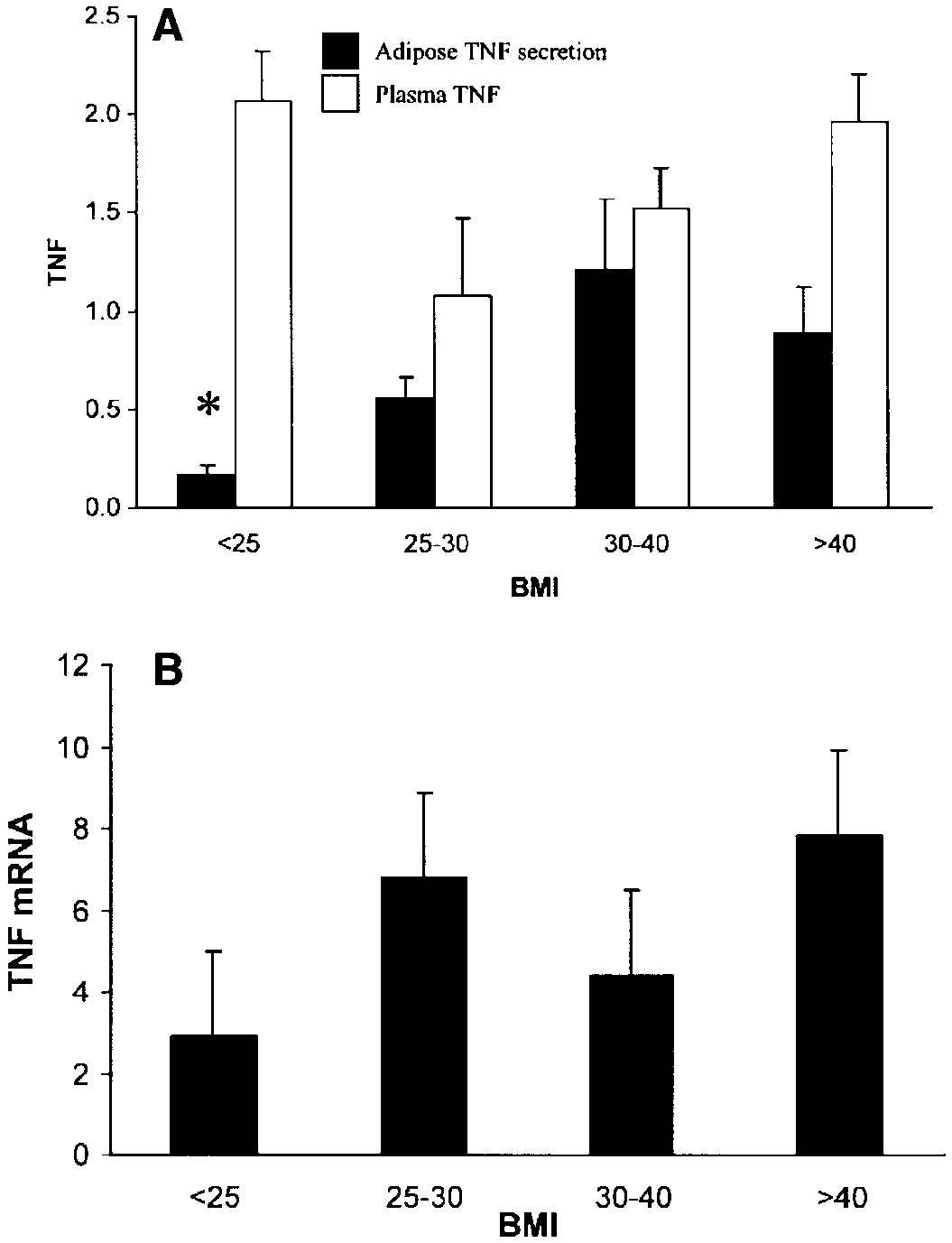 in subjects with a BMI between 30 and 40 kg/m2 (P Ͻ0.05). Subjects with a BMI Ͼ45 kg/m2 demonstratedslightly lower TNF secretion (0.90 Ϯ 0.21 pg/g DNA),but this was not significantly decreased compared withsubjects with a BMI of 30–40 kg/m2. This effect of BMIon TNF secretion was still present when women andCaucasians were each considered separately and whensubjects with impaired glucose tolerance were elimi-nated. TNF secretion from adipose tissue was also lowin subjects with low body fat. TNF secretion in subjectswith Ͻ30%, 30–45%, and Ͼ45% body fat was 0.16 Ϯ0.07 (n ϭ 10), 0.76 Ϯ 0.16 (n ϭ 14), and 1.1 Ϯ 0.28pg/g DNA (n ϭ 18, P Ͻ 0.05 vs. Ͻ30% group).
in subjects with a BMI between 30 and 40 kg/m2 (P Ͻ0.05). Subjects with a BMI Ͼ45 kg/m2 demonstratedslightly lower TNF secretion (0.90 Ϯ 0.21 pg/g DNA),but this was not significantly decreased compared withsubjects with a BMI of 30–40 kg/m2. This effect of BMIon TNF secretion was still present when women andCaucasians were each considered separately and whensubjects with impaired glucose tolerance were elimi-nated. TNF secretion from adipose tissue was also lowin subjects with low body fat. TNF secretion in subjectswith Ͻ30%, 30–45%, and Ͼ45% body fat was 0.16 Ϯ0.07 (n ϭ 10), 0.76 Ϯ 0.16 (n ϭ 14), and 1.1 Ϯ 0.28pg/g DNA (n ϭ 18, P Ͻ 0.05 vs. Ͻ30% group).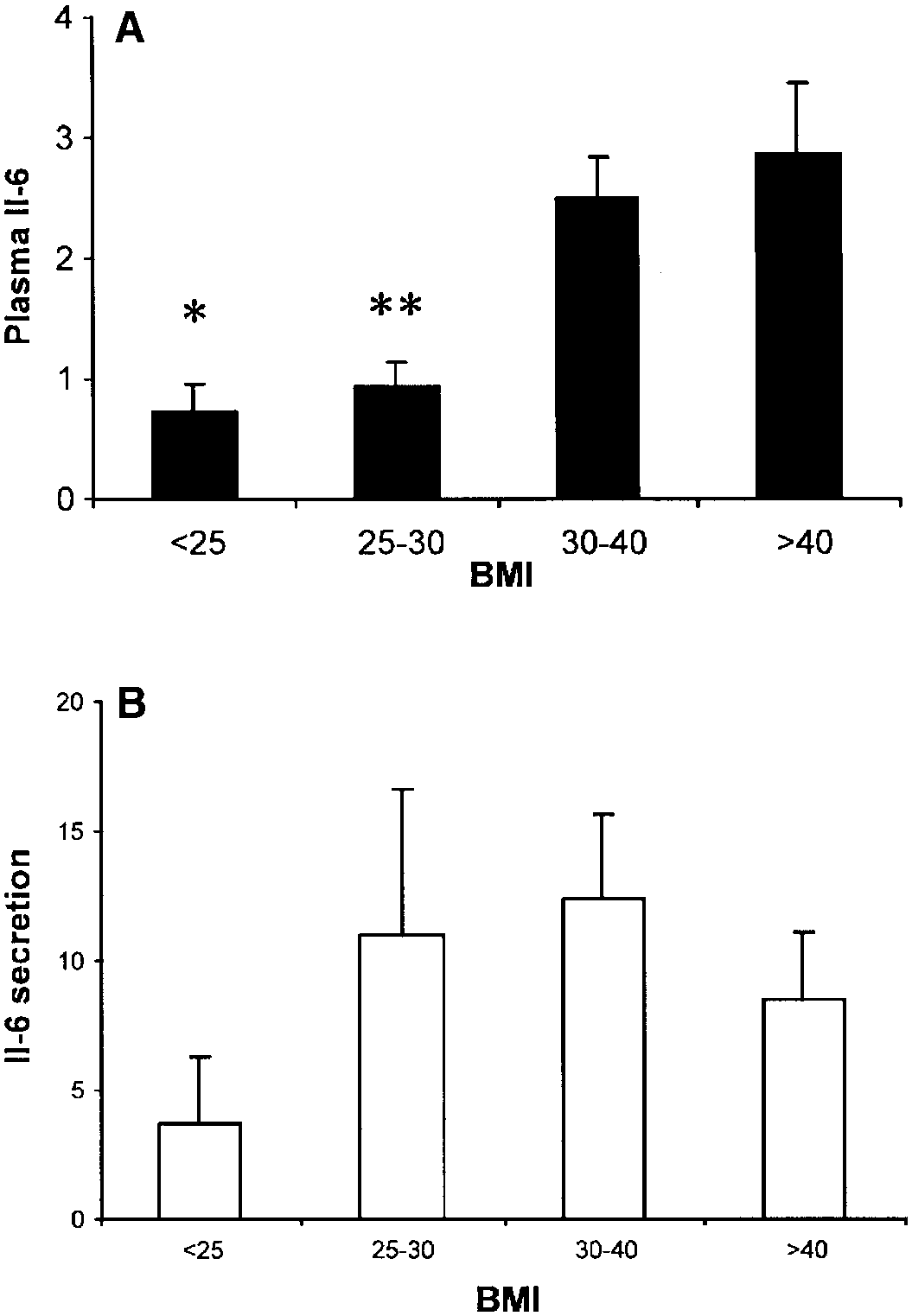
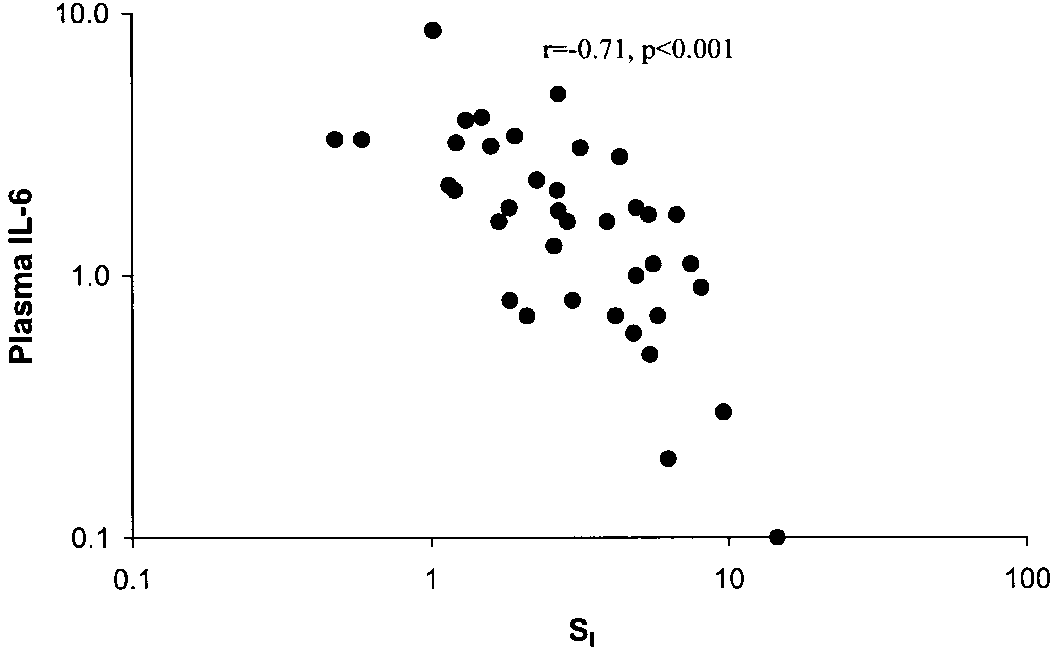
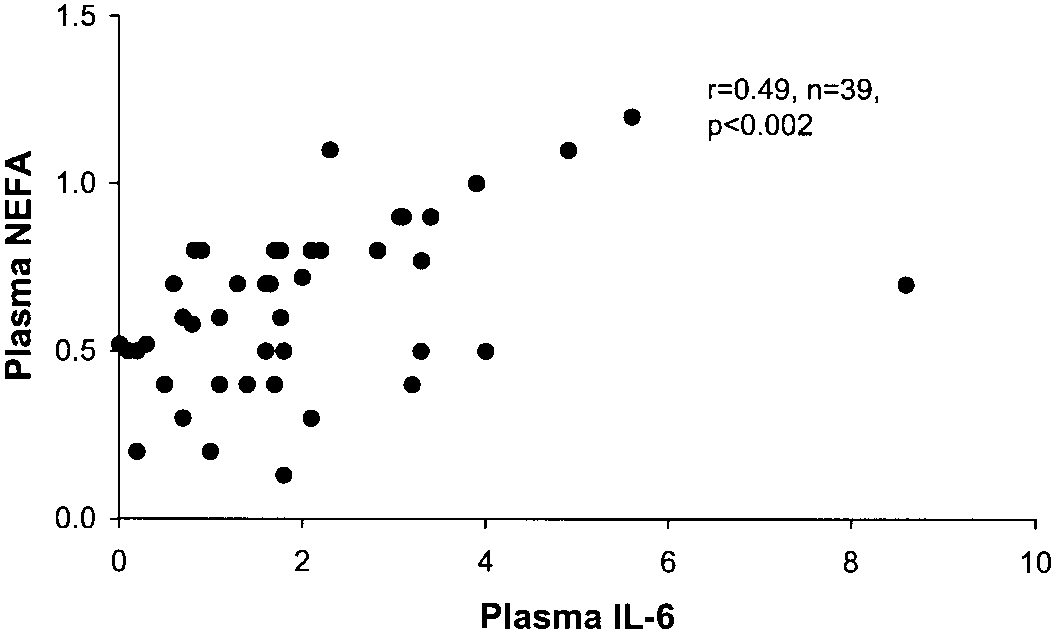 Fig. 5. Relationship between plasma IL-6 and SI. Plasma IL-6 wasmeasured as described in METHODS and expressed as pg/ml. Data arelog transformed.
Fig. 5. Relationship between plasma IL-6 and SI. Plasma IL-6 wasmeasured as described in METHODS and expressed as pg/ml. Data arelog transformed.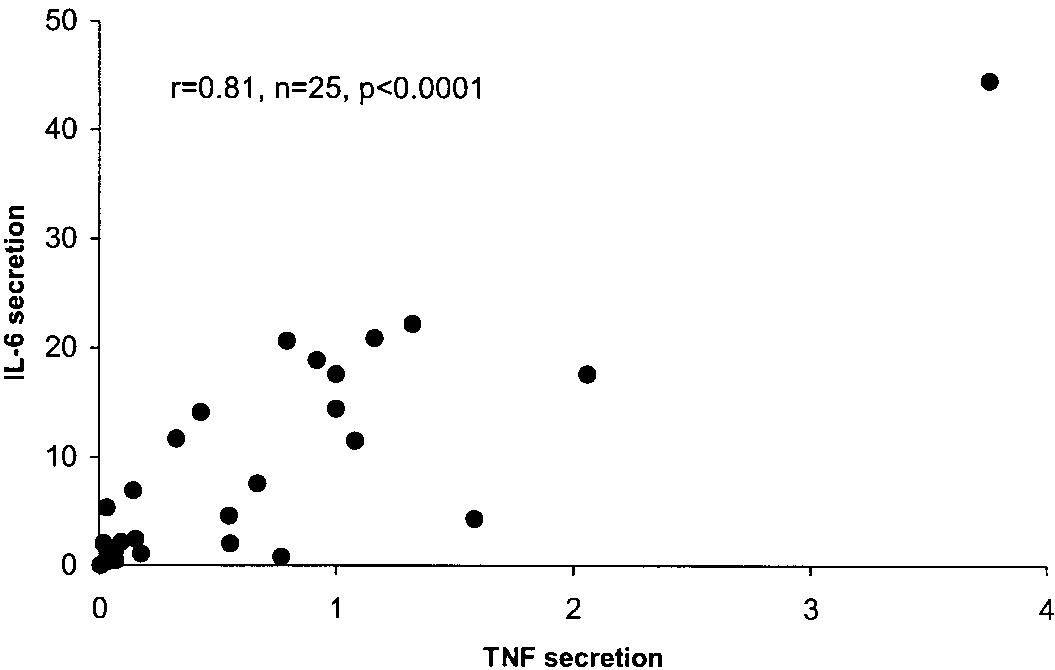 Table 2. Insulin-sensitive and insulin-resistant subjects matched for BMI and age
Values are means Ϯ SE. SI, insulin sensitivity; TNF, tumor necrosis factor; IL-6, interleukin-6. * P Ͻ 0.05 vs. insulin sensitive.
Table 2. Insulin-sensitive and insulin-resistant subjects matched for BMI and age
Values are means Ϯ SE. SI, insulin sensitivity; TNF, tumor necrosis factor; IL-6, interleukin-6. * P Ͻ 0.05 vs. insulin sensitive.